Методами многомерного анализа данных проведено сравнение формы и размеров листьев Alnus incana s.str., A. glutinosa, A. barbata и A. kolaensis при помощи классической и геометрической морфометрии. Мы выяснили, что: (1) результаты анализа данных, полученных методами геометрической и классической морфометрии, различаются; (2) A. kolaensis не следует рассматривать как таксон, обособленный от A. incana; (3) A. barbata представляет собой таксон видового ранга, наиболее близкий к A. glutinosa; (4) для описания формы листа методами геометрической морфометрии больше всего подходит разметка на основе концевых точек жилок второго порядка.
Род Alnus Mill. включает 30−40 видов, распространенных в умеренных широтах Северного полушария (Комаров, 1936; Давидов, 1972; Furlow, 1979; Цвелев, 2004). Некоторые виды заходят также в Южную Америку, а в Азии доходят до Бенгалии и Северного Вьетнама, но встречаются там только в горах. На севере отдельные виды доходят до лесотундры и тундры, а в горах поднимаются до субальпийского пояса. Большинством систематиков (Черепанов, 1955; Цвелев, 2002) род Alnus разделяется на несколько секций, среди которых следует назвать секцию Alnobetula Koch (часто выделяется в отдельный род Dushekia Opiz), секцию Alnus, к которой принадлежит, например, ольха клейкая — Alnus glutinosa (L.) Gaertn. и секцию Proskeimostemon Czer., которая распространена преимущественно в Европе и включает ольху серую — A. incana (L.) Moench. Следует отметить, однако, что данные анализа последовательностей ДНК (Navarro et al., 2003) не полностью согласуются с указанной системой.
Alnus incana — самый распространенный вид ольхи Евразии. Широко известен ее полиморфизм, причем существуют формы, которые часто считают отдельными видами, например, Alnus kolaensis Orlova (Орлова, 1954), произрастающая на юго-востоке Фенноскандии, в Норвегии, на Кольском полуострове и Карельском берегу Белого моря. Систематики по-разному понимают статус кольской ольхи. Так, Н.И. Орлова (1954), М.Л. Раменская и В.Н. Андреева (1982) считают ее отдельным видом; А.К. Скворцов (1959) и O. Nilsson (2000) — подвидом A. incana; многие авторы (Walters, 1993; Соколов, Филин, 1996) вовсе не выделяют этот таксон; а некоторые Н.Н. Цвелев (2002, 2004) авторы предполагает ее гибридное происхождение от скрещивания ольхи серой и клейкой, хотя в этом случае неясны ее взаимоотношения с давно описанным гибридом A. x pubescens Taush. Со своей стороны, O. Nilsson (2000) рассматривает A. x pubescens и A. kolaensis как отдельные формы. Дело осложняется тем, что в настоящее время на большей части ареала ольхи кольской не встречается один из предполагаемых “родителей” — A. glutinosa. Наконец, Е.В. Банаев и М.А. Шемберг (2000) считают, что некоторые экземпляры сибирской A. hirsuta (Spach.) Turcz. ex Rupr. неотличимы от A. kolaensis. Подобные формы свойственны и A. glutinosa, например, произрастающая на Кавказе ольха бородатая (A. barbata C.A. Mey.), таксономический статус которой колеблется от подвида до вида (Черепанов, 1995). Основные отличия этих форм ольхи указаны в таблице 1, составленной на основе литературных данных.
|
вид |
Alnus barbata |
Alnus cordata |
Alnus glutinosa |
Alnus incana |
Alnus incana var. argentata |
Alnus kolaensis |
Alnus x pubescens |
Alnus viridis |
|
жизненная форма |
Высокое дерево |
Дерево до 9-15 м |
Дерево 20-35 м, реже куст |
Дерево до 20-30 м или куст |
Дерево или куст |
Куст или дерево до 8 м |
Дерево или куст |
Куст до 3-4 м |
|
почки |
На ножке 2-7 мм, с 2 чешуями, клейкие |
На ножке 1-7 мм, с 2-4 чешуями, голые |
На ножке 2-7 мм, с 2 чешуями, клейкие |
На ножке 1-4 мм, с 2 чешуями, голые или опушенные |
На ножке 1-4 мм, с 2 чешуями, голые или опушенные |
На ножке 1-4 мм, с 2 чешуями, голые |
Промежуточные между A. incana и A. glutinosa |
Почти сидячие, с 3-6 черепитчатыми чешуями |
|
молодые побеги |
Железистые, кора коричневая |
Железистые, кора блестящая, коричневатая |
Железистые, кора зеленовато- или красно-коричневая |
Опушенные до войлочного опушения, кора серо-коричневая |
Опушенные до войлочного опушения, кора темно-серая |
Опушенные, кора красновато-коричневая или желтоватая |
Опушенные |
Железистые, кора зеленая или красновато-коричневая |
|
размер листьев |
6-10 см длиной и 5-8 см шириной |
2-12 см длиной |
4-10 см длиной и 2,5-8,5 см шириной |
(2,5)3,5-9 см длиной и (2,5)3,2-6 см шириной |
5,5-7,5(9) см длиной и 4,5-6(8) см шириной |
3-5(6) см длиной и 2,7-5 см шириной |
Более 6 см длиной |
2-7 см длиной и 2-4,5 см шириной |
|
форма листьев |
Овальные или более вытянутые, с округлым основанием и притупленной, иногда выемчатой верхушкой |
Овальные, с сердцевидным основанием и оттянутой тупоконечной верхушкой |
Обратнояйцевидные, реже овальные, с округлым основанием и выемчатой верхушкой |
Овально-ланцетная до округлой, основание округлое, верхушка заостренная |
В основном овальные, с округлым основанием и тупой или заостренной верхушкой |
Овальные или яйцевидные, основание округлое, верхушка обычно тупая |
Овальные, основание округлое, верхушка коротко заостренная |
Овальные, основание клиновидное, верхушка треугольная |
|
форма края листа |
Городчато-пильчатый |
Редко- и мелкозубчатый |
Городчато-пильчатый |
Дваждыпильчатый |
Дваждыпильчатый с крупными зубцами |
Дваждыпильчатый |
Городчато-пильчатый |
Слабо зубчатый |
|
нижняя сторона листа |
Светло-зеленая, опушенная в углах жилок и по жилкам |
Бледно-зеленая, голая (у молодых листьев пучки волосков в углах жилок) |
Зеленоватая, с редкими волосками по жилкам |
Серо-зеленая, опушенная |
Серо-зеленая, густо опушенная |
Серо-зеленые, по жилкам опушенные |
Сероватая, опушенная в углах жилок и по жилкам |
Зеленая, у молодых листьев клейкая |
|
количество жилок |
5-7(9) |
От 5 до 8 |
5-7(9) |
(5)7-11(13) |
(5)7-11(13) |
(5)6-8 |
От 5 до 11 |
От 4 до 8 |
|
зрелые соплодия |
6-8 мм шириной и 16-20 мм длиной |
10-15 мм шириной и 25-30 мм длиной |
(8)10-12 мм шириной и (7)9-14(17) мм длиной, на ножках до 20 мм |
10-17 мм длиной и 8-12 мм шириной, сидячие или на ножках до 6 мм |
10-17 мм длиной и 8-12 мм шириной, сидячие или на ножках до 6 мм |
До 10 мм шириной |
10-17 мм длиной и 8-12 мм шириной, на коротких ножках |
8-15 мм длиной и 3-5 мм шириной, на ножках до 24 мм |
|
орешек |
Округлый, 2,5-4 мм длиной и 2,2-3,2 мм шириной, крыло 0,3-1,2 мм шириной |
С крылом |
Округлый, 2,5-4 мм длиной и 2,2-3,2 мм шириной, крыло 0,3-1,2 мм шириной |
Эллиптический, 3-4,5 мм шириной и 2-3,8(4) мм длиной, крыло 0,5-1,5 мм шириной |
Эллиптический, 3-4,5 мм шириной и 2-3,8(4) мм длиной, крыло 0,5-1,5 мм шириной |
Эллиптический, крыло 1-1,5 мм шириной |
Промежуточный между A. incana и A. glutinosa |
Орешек с пленчатым крылом |
К сожалению, подробный таксономический анализ вышеназванных форм до сих пор не проводился. Для такого анализа можно использовать кариологические данные, данные классической морфометрии (Банаев, Шемберг, 2000), анализ последовательностей ДНК (King et al., 1998; Navarro et al., 2003), а также достаточно новый метод, появившийся в девяностых годах прошлого века — геометрическую морфометрию (Павлинов, 2002), который хорошо зарекомендовал себя при изучении таксономического разнообразия сложных групп, включающих таксоны гибридного происхождения (Jensen et al., 2002; Shipunov, Bateman, in press).
Для сравнения различных видов ольхи Е.В. Банаев и М.А. Шемберг (2000) применяли методы классической морфометрии, причем из 17 исследованных характеристик 11 являлись характеристиками листовой пластинки, которые, по мнению авторов, имеют большое таксономическое значение. В этой связи мы поставили задачу провести морфологический анализ признаков листьев видов ольхи Европейской России для того, чтобы: (1) оценить морфологическое разнообразие и возможные таксономические отношения, а также (2) сравнить результаты, полученные методами классической и геометрической морфометрии.
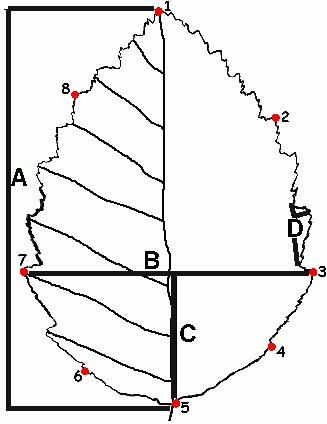
Сбор материала проводили в 2002−2004 гг. в Удомельском и Вышневолоцком районах Тверской области на биостанции Молдино, в Лоухском районе республики Карелия, на Соловецких островах Белого моря, в западном Закавказье (район Большого Сочи), а также ботанических садах Москвы и Лондона. Кроме того, изучены гербарные коллекции Московского государственного университета им. М.В. Ломоносова (МГУ) (MW), Гербария Ботанического Сада (ГБС) РАН (MHA) и Биологического Института им. Комарова (БИН) РАН (LE). Всего было исследовано 785 образцов из 171 местообитания. Для измерений с каждого растения выбирался наибольший лист листовой серии удлиненного побега текущего года. Были сделаны промеры длины и ширины листовой пластинки, расстояния от ее основания до положения наибольшей ширины, количества жилок первого порядка с левой стороны листа, размера наибольшего зубца первого порядка (рис. 1), а также опушение листа и побега по трехбалльной шкале. Изображения всех измеренных листьев (или только их контуры с отметками мест выхода жилок) были переведены в цифровой вид при помощи планшетного сканера (разрешение 200 dpi, масштаб 100%) и сохранены в файлах графических форматов PCX или PNG (для отсканированных контуров) или JPEG (для отсканированных листьев).
A – длина листа, B – ширина листа, C – расстояние от черешка до наибольшей ширины листа, D – длина зубца первого порядка
Для характеристики формы листа мы использовали подход, связанный с расстановкой так называемых меток (“landmarks”, Павлинов, 2002; Adams et al., 2003). Выбор соответствующих точек оказался непростой задачей, поскольку листья геометрически довольно однородны и лишь верхушка и основание листа хорошо подходят на роль меток. Двух меток, конечно, совершенно недостаточно, и для расстановки большего числа меток мы применяли два различных подхода. Во-первых, мы ставили 8 меток в точках наибольшей кривизны контура листа, считая основание и верхушку (рис. 1). Метки, полученные таким образом, хорошо характеризуют форму листа, однако нельзя утверждать с уверенностью, что они гомологичны для разных листьев; речь может идти лишь о “геометрической гомологии” (Павлинов, 2002). Другой подход состоял в расстановке 12 меток: на пересечениях первых пяти пар (считая от основания листа) жилок первого порядка с контуром листа, а также у основания и верхушки листа. Такой подход к описанию формы листа первые предложен R. Jensen с соавторами (2002), однако, насколько нам известно, еще ни разу не использовался. 12 меток также неплохо характеризуют форму листа, но если лист имеет больше 5 пар жилок, то верхняя половина листа оказывается “свободной” от меток, что снижает точность описания формы. Таким образом, оба метода расстановки меток имеют свои преимущества и недостатки.
Координаты точек снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig (Rohlf, 2004a). Исходные координаты экземпляров нормировали относительно так называемой эталонной (усредненной, или консенсусной) конфигурации при помощи прокрустова наложения. Координаты эталонной конфигурации, а также значения главных, относительных и частных трансформаций, характеризующие меру отличия образца от эталона, вычисляли при помощи программы tpsRelw (Rohlf, 2004b). Редактирование и конвертирование файлов данных производили при помощи вспомогательной программы tpsUtil (Rohlf, 2004c)
Для анализа данных классической морфометрии, а также для обработки выходных данных программы tpsRelw мы применяли многомерные статистические методы, такие как анализ главных компонент, кластерный и дискриминантный анализ. Последний вид анализа использовался нами для сравнения различных классификаций исходных форм. Для этого вычислялись так называемые “ошибки классификации” (misclassification errors), которые являются мерой несоответствия между заранее заданной классификацией и распределением форм по группам, полученным в результате анализа данных. Для всех статистических расчетов использовалась компьютерная статистическая среда R (R Development Core Team, 2004).
Поскольку нас преимущественно интересовали взаимоотношения с близкими видами ольхи двух относительно далеких друг от друга форм (A. kolaensis и A. barbata), все образцы были разделены на две пересекающиеся группы, в каждой из которых все перечисленные этапы анализа производились независимо. В состав первой группы вошли природные и гербарные образцы A. incana (только из центральной и северной России), [A. incana var. argentata Norrl. (= A. argentata (Norrl.)) Tzvel.], [A. x pubescens], A. kolaensis (= A. incana subsp. kolaensis (Orlova) A. Love et D. Love, = A. incana var. virescens Wahlenb.), A. glutinosa (только из центральной и северной России), [A. viridis (Chaix) DC.] и [A. cordata Loisel.]. Виды, названия которых заключены в квадратные скобки, не вошли в анализ с использованием 12 меток. Последние два вида (относящиеся к секции Alnobetula) были взяты в качестве “якорных точек”, которые позволяют оценить таксономическое значение возможных различий. В состав второй группы вошли природные и гербарные образцы A. incana (только из центральной и южной России), A. barbata C.A. Mey., A. glutinosa (только из центральной и южной России), [A. x pubescens], [A. viridis] и [A. cordata].
Анализ главных компонент для данных классической морфологии показывает высокую полиморфность изученных групп ольхи по признакам листа. Вместе с тем на графике двух первых компонент (рис. 2) можно увидеть определенную структуру. Прежде всего, хорошо выделяется три “облака”, соответствующие A. glutinosa, A. viridis и формам, близким к A. incana. Последняя группа не имеет четкой структуры, образцы из природных популяций Карелии значительно перекрываются с гербарными образцами A. kolaensis и образцами A. incana. Гибриды A. x pubescens также находятся внутри этого “облака”. Некоторые образцы A. kolaensis “проникают” внутрь A. glutinosa. Парадоксально, что великолепно отличающаяся по форме листьев A. cordata не занимает обособленного положения на графике, а располагается внутри “облака” A. glutinosa.
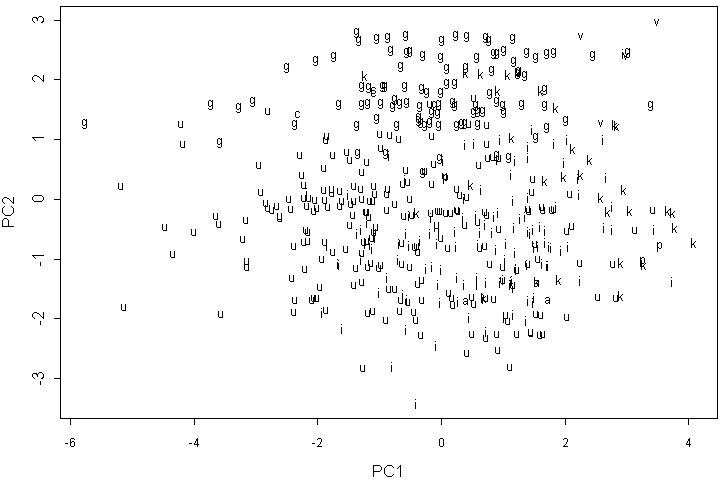
Буквами латинского алфавита обозначена видовая принадлежность образцов: a — A. incana var. argentata, c — A. cordata, g — A. glutinosa, i — A. incana, k — A. kolaensis, p — A. x pubescens, u — образцы A. incana s.l. из природных популяций северной Карелии и Соловецких островов, v — A. viridis.
На графике первых двух компонент для данных геометрической морфометрии по 8 меткам мы видим три очень четких “облака” (рис. 3). Первое образовано A. glutinosa, второе образовано формами, близкими к A. incana, а третье — образцами A. cordata. Внутри второго “облака” наблюдается сильное перекрытие всех форм. Многие образцы A. kolaensis занимают периферийное положение, в основном ближе к A. glutinosa. Этот тип анализа не привел к выделению отдельного “облака” для A. viridis.
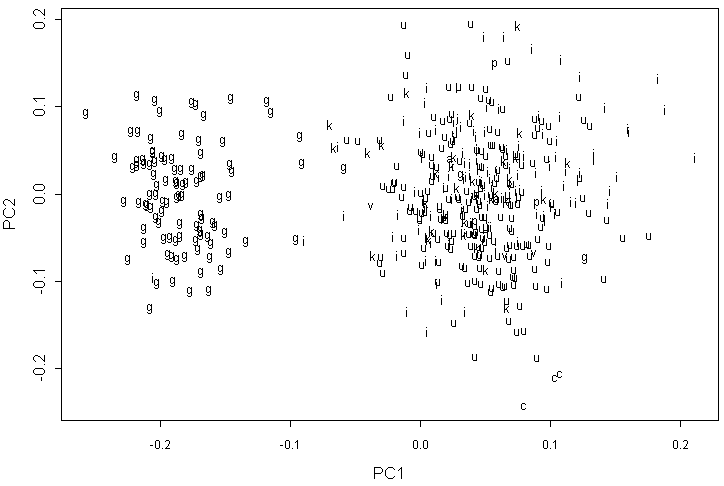
Обозначения как на рис. 2.
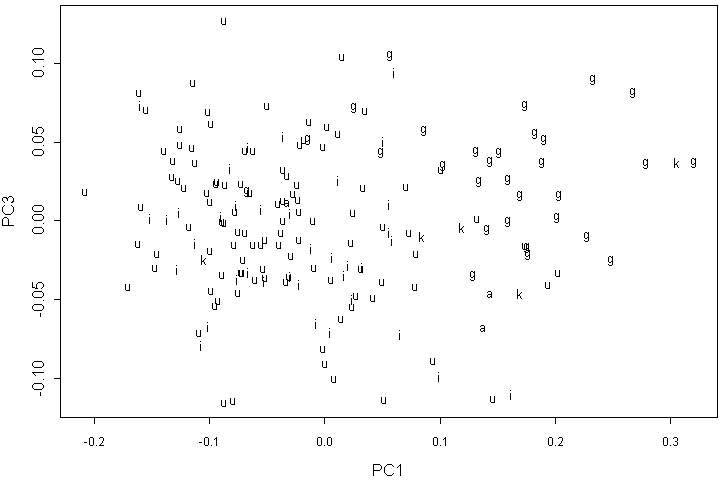
В анализе данных геометрической морфометрии по 12 меткам второй и третий компоненты имеют приблизительно равные вклады, причем наиболее четкое распределение форм наблюдается на графике первой и третьей компонент (рис. 4). Хорошо заметны “облака” для A. glutinosa и форм, близких к A. incana. Некоторые образцы из природных популяций Карелии и A. kolaensis занимают периферическое положение во второй группе, а в некоторых случаях даже попадают внутрь группы A. glutinosa.
Для сравнения классификаций были выбраны четыре гипотезы (табл. 2), которые отражают различные подходы к таксономической принадлежности образцов из северной Карелии и к самостоятельности A. kolaensis. Налицо значительное снижение ошибок классификации в случае принятия четвертой гипотезы.
|
Гипотеза |
Средний процент ошибок классификации |
|
|
Геометрическая морфометрия |
Классическая морфометрия |
|
|
Alnus kolaensis, A. incana s.str., A. incana var. argentata и формы из северной Карелии — отдельные группы |
26.3% |
48.8% |
|
То же, но формы из северной Карелии принадлежат к A. kolaensis |
22.4% |
39.8% |
|
То же, но формы из северной Карелии принадлежат к A. incana s.str. |
20.5% |
47.6% |
|
Все перечисленные формы принадлежат к одной группе |
3.7% |
31.1% |
Анализ главных компонент для данных классической морфологии показывает четкую структуру (рис. 5). Видны три обширные группы: A. incana, A. glutinosa и A. barbata. Alnus viridis (в отличие от A. cordata) образует более или менее четкую группу в верхнем левом углу графика. Интересно, что в отсутствие других форм A. incana два образца A. x pubescens группируются вместе и достаточно далеко от A. barbata.
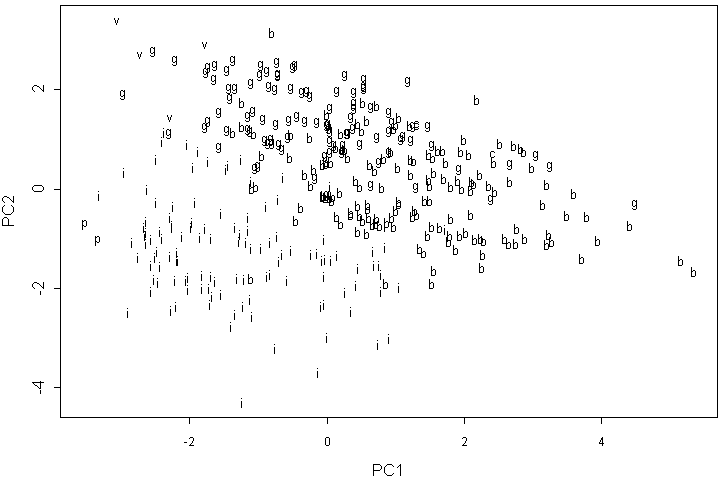
Буквами латинского алфавита обозначена видовая принадлежность образцов: b — Alnus barbata, c — A. cordata, g — A. glutinosa, i — A. incana, p — A. x pubescens, v — A. viridis.
В анализе данных геометрической морфометрии по 8 меткам второй и третий компоненты имеют приблизительно равные вклады, причем наиболее четкое распределение форм наблюдается на графике первой и третьей компонент (рис. 6). Видны два резко отличающихся “облака”, причем образцы A. barbata находятся преимущественно среди A. incana, хотя некоторые формы попали к A. glutinosa. Все прочие формы (в том числе A. viridis и A. cordata) не занимают обособленного положения.
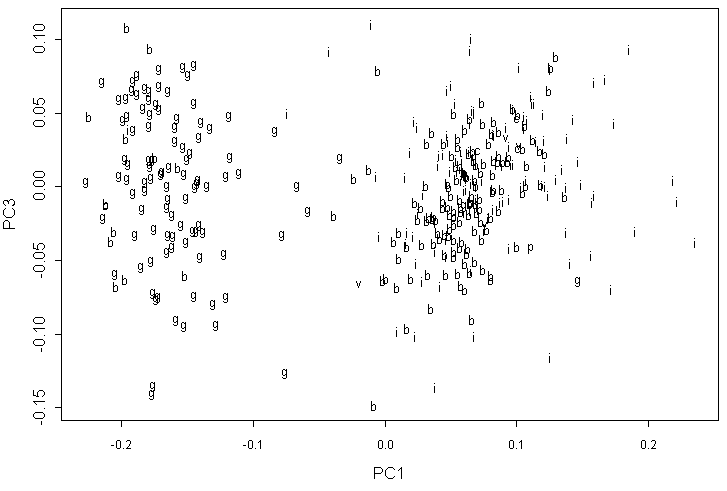
Обозначения как на рис. 5.
На графике первых двух компонент для данных геометрической морфометрии по 12 меткам (рис. 7) структура выражена значительно меньше, причем в этом случае образцы A. barbata не перекрываются с A. incana и занимают более или менее промежуточное положение между этим последним видом и A. glutinosa (со значительным перекрытием).
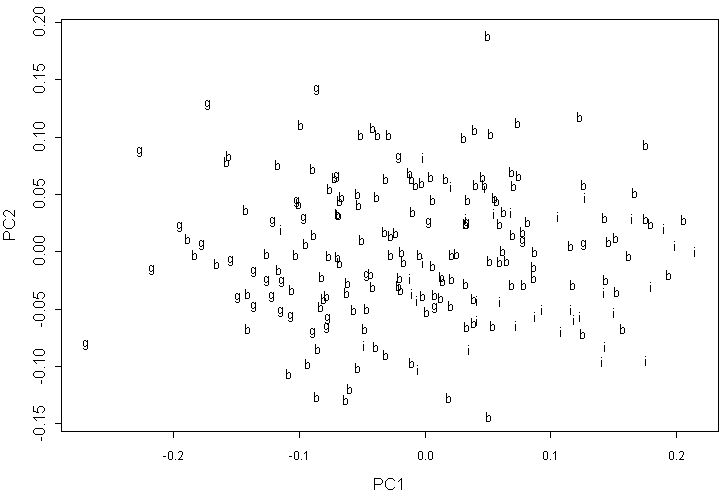
Обозначения как на рис. 5.
Для сравнения классификаций были выбраны три гипотезы (табл. 3). Характерно увеличение ошибок классификации при принятии любой гипотезы, объединяющей A. barbata с другими видами.
|
Гипотеза |
Средний процент ошибок классификации |
|
|
Геометрическая морфометрия |
Классическая морфометрия |
|
|
Alnus barbata отличается от A. glutinosa и A. incana |
15.6% |
9.2% |
|
Alnus barbata и A. incana относятся к одной группе |
17% |
13.9% |
|
Alnus barbata и A. glutinosa относятся к одной группе |
17% |
19.6% |
Многомерный анализ данных классической морфометрии свидетельствуют о том, что формы, близкие к A. incana, не обнаруживают обособленности ни друг от друга, ни от A. incana s.str. Лишь гербарные образцы A. kolaёnsis несколько обособлены от остальных форм, хотя и здесь нельзя провести четкой границы между ними и природными образцами из Карелии. Это и неудивительно, поскольку A. kolaёnsis выделялась преимущественно по размерным признакам листьев. Наши личные наблюдения показывают, что листья, сходные с “типичной” А. kolaёnsis, можно найти как на укороченных побегах, так и на угнетенных удлиненных побегах A. incana из северной Карелии. Некоторые формы, в особенности растущие по окраинам сфагновых болот, имеют листья в основном “kolaёnsis-типа”. Все это может свидетельствовать об экологической приуроченности мелколистных форм. Близость обоих образцов A. x pubescens к A. kolaёnsis не позволяет на основании размерных данных отбрасывать гипотезу о гибридном происхождении A. kolaёnsis. Неспособность данного анализа выделить A. cordata показывает различие между классической и геометрической морфометрией: в то время как A. cordata легко выделяется по форме листа (см. следующий анализ), размерные признаки листа этого вида не отличают ее от A. glutinosa.
Резкое различие между A. glutinosa и всеми остальными формами в анализе по 8 точкам объясняется, на наш взгляд, не только большей информативностью признаков геометрической морфометрии, но и типом постановки меток, при котором вторая и восьмая точки для большинства образцов ольхи клейкой были сильно смещены к верхушке за счет наличия выемки. Таким образом, “левая” группа на графике маркирует листья с выемкой. Периферийное положение многих образцов A. kolaёnsis свидетельствует об отклонениях формы листа по сравнению с типичными A. incana. Alnus viridis не отличается принципиально по форме листа от A. incana, а образцы A. x pubescens не группируются с A. kolaёnsis: таким образом сходство между этими формами лишь размерное. Alnus incana var. argentata не отличается от типичных A. incana ни в одном из типов анализа.
Анализ с использованием 12 точек, в общем, соответствует предыдущему, но разница между группами выражена менее резко за счет того, что данный метод не учитывает выемку на верхушке листа. Таким образом, разница между первым и вторым анализом возникла не только за счет того, что классическая морфометрия не учитывает наличие выемки; очевидно, что форма листа в целом позволяет более четко распределить образцы по группам.
Результаты дискриминантного анализа свидетельствуют о том, что для анализируемых данных лучше всего принять гипотезу о единстве всех форм, близких к A. incana. Об этом же говорит и сравнение с “якорными точками”: ни в одном случае различия между формами A. incana не велики так, как велики они между “нормальными” видами. Наконец, анализ показывает, что образцы из северной Карелии и с Соловецких островов также должны быть отнесены к A. incana.
Alnus barbata и близкие виды
Обращает на себя внимание различие между всеми тремя видами анализа для этой группы: в то время как анализ признаков классической морфометрии выделяет A. barbata в отдельное “облако”, методы геометрической морфометрии сближают ее с A. incana (8 точек) или с A. glutinosa (12 точек). Видно, что в данном случае классическая морфометрия несколько лучше отражает разнообразие исходных данных (об этом свидетельствуют и данные дискриминантного анализа), а на данные, полученные в анализе по 8 точкам, влияет наличие или отсутствие выемки у листьев A. barbata (возможны оба варианта, что и приводит к “миграции” части образцов в группу A. glutinosa). Данные по 12 точкам свидетельствуют от том, что по форме листа A. barbata ближе всего к A. glutinosa; об этом говорят и другие признаки этих видов (см. табл. 2).
Дискриминантный анализ дает довольно неожиданный результат: процент ошибок классификации при уменьшении числа групп возрастает. Возрастание процента ошибок однозначно говорит за принятие такой таксономической гипотезы, в которой все три группы рассматриваются как отдельные таксоны. К сожалению, провести сравнение с остальными видами не удается, поскольку “якорные” группы ни в одном из случаев четко не выделяются. В общем, наши данные говорят о том, что A. barbata — отдельный таксон видового ранга. Следует отметить также, что наиболее близким таксоном к A. barbata является A. glutinosa, причем вряд ли можно говорить о гибридной природе первого таксона.
Общие выводы
Таким образом, полученные результаты говорят за то, чтобы рассматривать A. kolaёnsis в составе A. incana (как разновидность или даже как экологическую форму), а A. barbata — как отдельный вид, близкий к A. glutinosa.
Наши данные свидетельствуют также о том, что результаты анализа данных, полученных методами геометрической и классической морфометрии, могут значительно различаться. Эта ситуация отличается от наблюдавшейся при анализе формы губы цветка Dactylorhiza (Shipunov, Bateman, in press) и листьев Acer (Jensen et al., 2002). Тест Мантеля, проведенный нами для выяснения соответствия (корреляции) между методами классической и геометрической морфометрии, свидетельствует о незначительном соответствии (r=0.1008, p=0.01, 1000 репликаций). По-видимому, различия состоят не только в способности или неспособности отразить характерный признак наличия выемки, но значат также, что классическая и геометрическая морфометрия описывают существенно различающиеся аспекты морфологии листьев ольхи. Таким образом, эти два типа данных могут использоваться совместно. Что же касается различных способов размещения меток, то наш анализ свидетельствует о преимуществах подхода с использованием концевых точек жилок второго порядка (метод 12 точек).
Благодарности
Сбор большей части данных для настоящей работы проводился в рамках биологических практик Московской Гимназии на Юго-западе N1543. Мы благодарим всех участников и руководителей практик за помощь, оказанную нам в сборе и подготовке материала. Мы также выражаем благодарность сотрудникам гербариев МГУ (MW), ГБС (MHA) и БИН РАН (LE) за содействие в работе.
Список литературы
Банаев Е. В., Шемберг М. А. Ольха в Сибири и на Дальнем Востоке России // Новосибирск, 2000. 100 с.
Давидов М. В. Лесоводственно-биологическая характеристика ольхи // М., 1972. С. 4−13.
Комаров В. Л. Род Ольха – Alnus Gaertn. // Флора СССР. М.;Л., 1936. С. 306−319.
Орлова Н. И. Новый вид ольхи с Кольского полуострова // Бот. мат. Герб. БИН. 1954. Т. 16. С. 70−75.
Павлинов И.Я., Микешина Н.Г. Принципы и методы геометрической морфометрии // Журн. общ. биол. 2002. Т. 63. N 6. С. 473−493.
Раменская М. Л., Андреева В. Н. Alnus Mill. — Ольха // Определитель высших растений Мурманской области и Карелии. Л., 1982. С. 169−170.
Скворцов А. К. Новые флористические находки в районе Денежкина Камня (Северный Урал) // Бот. мат. Герб. БИН. 1959. Т. 19. С. 558−571.
Соколов Д. Д., Филин В. Р. Alnus Mill. — Ольха // Определитель сосудистых растений окрестностей ББС МГУ. М., 1996. С. 76.
Цвелев Н.Н. О родах Betula L. и Alnus Mill. (Betulaceae) в Восточной Европе. // Новости систематики высших растений. Т. 34. Спб., 2002. C. 47–70.
Цвелев Н.Н. Ольха — Alnus Mill. // Флора Восточной Европы. Т. 11. Спб., 2004. С. 87–90.
Черепанов С. К. Система рода Alnus Mill. s. str. и близких к нему родов // Бот. мат. герб. БИН. М.;Л., 1955. Т. 17. С. 91−105.
Черепанов С. К. Сосудистые растения России и сопредельных государств. Спб., 1995. 990 с.
Adams D.C., Rohlf F.J., Slice D.E. Geometric morphometrics: ten years of progress following the “revolution”. Ecology and Evolution, SUNY at Stony Brook. 2002.
Furlow J. J. The systematic of American species of Alnus (Betulaceae) // Rhodora. Vol. 81. N. 825. 1979. P. 1−69.
Jensen R.J., Ciofani K.M., Miramontes L.C. Lines, outlines and landmarks: morphometric analyses of leaves of Acer rubrum, Acer saccharinum (Aceraceae) and their hybrid // Taxon. 2002. V. 51, N 3. P. 475–492.
King A. R., Ferris C. Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn. // Molecular ecology. 1998. Vol. 7. P. 1151−1161.
Navarro E., J. Bousquet, A. Moiroud, A. Munive, D. Piou, Ph. Normand. Molecular phylogeny of Alnus (Betulaceae), inferred from nuclear ribosomal DNA ITS sequences // Plant and Soil. Vol. 245. P. 207−217. 2003.
Nilsson O. Alnus Mill. // Flora Nordica. Stokholm, 2000. Vol. 1. P. 203−208.
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. 2004.
Rohlf F.J. tpsDig. N.Y.: State Univ. at Stony Brook. Version 1.40. 2004a. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph
Rohlf F.J. tpsRelw: relative warps. N.Y.: State Univ. at Stony Brook. Version 1.39. 2004.и [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph
Rohlf F.J. tpsUtil — TPS utility program. N.Y.: State Univ. at Stony Brook. Version 1.40. 2004с. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph
Shipunov A. B., R. M. Bateman. Geometric morphometrics as a tool for understanding Dactylorhiza (Orchidaceae) diversity in European Russia // Biol. J. Linn. Soc., in press.
Walters S. M. Alnus Mill. // Flora Europea. 2nd ed. Cambridge, 1993. Vol. 1. P. 68−70.