На территории России произрастает несколько видов рода плющ (Hedera). Многие виды этого рода плохо определяются. Это относится и к двум видам, произрастающим на Западном Кавказе: Hedera helix L. и H. colchica (K. Koch) K. Koch. Такая неопределенность связана прежде всего с большой изменчивостью листьев, по признакам которых обычно проводится определение (например, длина листа, количество лопастей и т.п.). Существуют и другие признаки, различающие виды. Это, в частности, число хромосом (у H. helix 2n=48, а у H. colchica 2n=192 - Vargas et al., 1999), запах (резкий и мускатный соответственно - Зернов, в печати), строение волосков (звездчатые и щитовидные соответственно - Ackerfield, 2000). Однако эти признаки в природе изучать трудно. Поэтому целью нашей работы было выявление оптимальных признаков, позволяющих классифицировать исследуемые растения на основании прежде всего метрических характеристик листьев.
Работа велась в рамках весенней биологической практики гимназии 1543 в районе города Туапсе и города Сочи Краснодарского края.
Измерения проводились на четырёх точках (площадках):
1) Каменный карьер. Сырой каштановый лес на юго-западном склоне двухсотметровой гряды.
2) Каштановый ручей. Русло ручья, впадающего в море с крутого обрыва и склоны, поросшие грабом и буком.
3) Водопад на реке Агой. Окрестности шестиметрового водопада в полутора километрах от посёлка Агой.
4) Барановское ущелье, по которому протекает река Восточный Дагомыс. В пятнадцати метрах от кромки воды, в сыром каштаново-самшитовом лесу.
Листья плющей разнообразны. Существует три рода побегов: побеги, на которых вырастают соцветия и впоследствии плоды (генеративные) побеги, вегетативные побеги, стелящиеся по земле (наземные) и вегетативные побеги, забирающиеся вверх по стволам деревьев (наствольные). Мы старались измерять все три типа побегов. Кроме этого, листья могут быть расположены не только на упомянутых (удлинённых), но и на укороченных побегах. Измерения листьев на последних не проводились. Но и это ещё не всё - на каждом побеге листья меняют свою форму и размеры, и происходит это вполне закономерно. Часть закономерно изменяющегося цикла мы, следуя Кренке (1933), назвали серией. Таким образом, измерения проводились по сериям - серией считался последовательно расположенный набор листьев, начинающийся с небольших листьев и заканчивающийся опять-таки набольшими. В качестве границы серии выбиралась граница годового прироста. Серия измерялась в направлении к верхушке побега.
Были выбраны следующие параметры измерения (в скобках приведены условные обозначения признаков, использовавшиеся для компьютерной обработки):
Таблица 1. Измеренные признаки.
| Код признака | Объяснение |
| BLANK.N | Номер бланка |
| PLOSCH.N | Номер площадки (см. выше) |
| TIP.POB | Тип побега (0 - наземный; 1 - наствольный; 2 - генеративный) |
| POBEG.N | Номер серии |
| N.L.SER | Порядковый номер листа в серии |
| L.MEZHD | Длина междоузлия, мм |
| L.CHER | Длина черешка, мм |
| W.LOC | Положение наибольшей ширины листа, мм |
| DIAG | Длина диагонали листа - расстояние от основания листа до точки перегиба в первой четверти его окружности, мм |
| L.MAX | Максимальная длина листа, мм |
| W.MAX | Максимальная ширина листа, мм |
| PARTS | Количество лопастей листа |
| WHO | Человек, измерявший листья |
| SMELL | Запах листьев (0 - слабый (мускатный), 1 - резкий) |
| TRICH | Вид волосков (1- звездчатые; 2- чешуевидные) |
Измерения проводились металлической линейкой. Для определения типа волосков использовался бинокулярный микроскоп. Всего был измерен 191 лист и 21 серия.
Данные были обработаны при помощи пакетов STATISTICA 5.0 (STATISTICA, 1999) и R (Ihaka, Gentleman, 1996).
Описательная статистика (см. табл. 2) говорит о том, что там, где величина стандартного отклонения низка по сравнению со средним значением, эта величина по абсолютному значению велика, и наоборот. Значит, исследуемые признаки очень изменчивы и поэтому по отдельности не очень подходят для классификации.
Таблица 2. Описательная статистика для используемых признаков.
| Valid N | Mean | Median | Minimum | Maximum | Lower Quartile | Upper Quartile | Std.Dev. | |
| BLANK.N | 191 | 6,36 | 6 | 1 | 13 | 4 | 9 | 3,20 |
| PLOSCH.N | 191 | 2,12 | 2 | 1 | 4 | 1 | 3 | 1,03 |
| TIP.POB | 191 | ,97 | 1 | 0 | 2 | 0 | 2 | ,84 |
| POBEG.N | 191 | 10,54 | 10 | 1 | 21 | 5 | 16 | 6,28 |
| N.L.SER | 191 | 6,20 | 5 | 1 | 20 | 3 | 8 | 4,38 |
| L.MEZHD | 190 | 33,29 | 26, | 0 | 160 | 19 | 40 | 25,65 |
| L.CHER | 191 | 34,92 | 29 | 2 | 163 | 15 | 45 | 27,79 |
| W.LOC | 189 | 12,67 | 10 | 0 | 55 | 5 | 17 | 10,70 |
| DIAG | 191 | 26,10 | 19 | 2 | 87 | 13 | 36 | 18,26 |
| L.MAX | 190 | 45,09 | 36, | 4 | 120 | 25 | 62 | 26,64 |
| W.MAX | 185 | 42,68 | 35 | 5 | 105 | 27 | 54 | 22,61 |
| PARTS | 186 | 2,73 | 3 | 1 | 5 | 1 | 3 | 1,29 |
| WHO | 191 | 4,83 | 4 | 1 | 8 | 4 | 6 | 2,02 |
| SMELL | 55 | ,64 | 1 | 0 | 1 | 0 | 1 | ,49 |
| TRICH | 191 | 1,28 | 1 | 1 | 2 | 1 | 2 | ,45 |
Кроме того, были использованы перекодированные признаки. Для перекодировки были использованы некоторые из первоначально измеренных признаков (см. табл. 3.)
Таблица 3. Перекодированные признаки.
| Признак | Как перекодировался | Объяснение |
| L.CH | L.MAX/L.CHER | Приподнятость листа над субстратом |
| L.Q | L.MAX/W.MAX | Форма листа: округлая, эллиптическая или ланцетовидная |
| L.VY | L.MAX/W.LOC | Форма листа: яйцевидность, эллиптичность или лопатчатость |
| L.W | DIAG/(sqrt(2W.MAX^2)) | Отношение диагонали к теоретической диагонали, вычисленной из расчета, что лист округлый |
Описательная статистика перекодированных признаков показывает меньщий разброс, и, стало быть большую пригодность данных признаков.
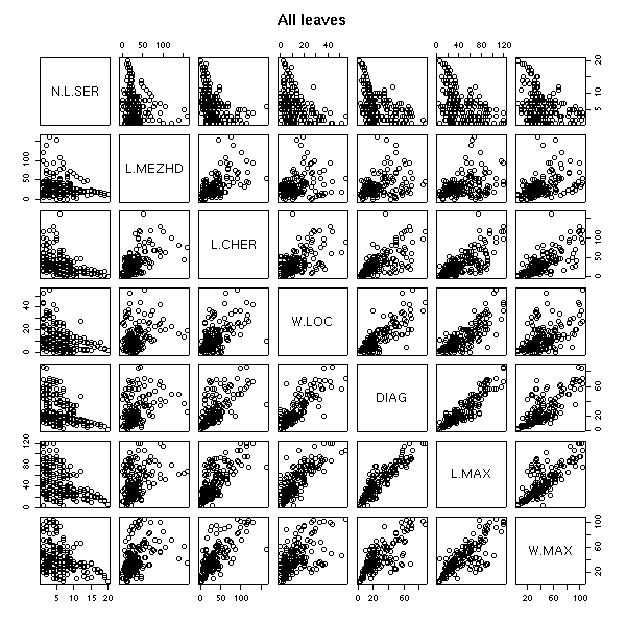
Анализ корреляций (табл. 4,5; рис. 1) говорит о том, что наибольшие корреляции наблюдаются между:
1) Длиной междоузлия, длиной черешка и всеми параметрами листа (колеблются в пределах 0,5-0,8). Из перекодированных признаков в эту группу входит только отношение диагоналей.
2) Количеством лопастей листа и остальными параметрами листа, в том числе с перекодированными признаками, за исключением приподнятости (около -0,5)
3) Количеством лопастей листа и типом волосков (-0,54)
4) Запахом и человеком, измерявшим плющи (0,9)
5) Всеми перекодированными признаками (около 0,5)
Видно, что запах - признак очень ненадежный, т.к. измерялся он не на всех растениях и во многих случаях вызывал затруднения в классификации, а корреляционный анализ показывает, что он субъективен.
Внимания заслуживает факт относительно низких корреляций измеренных признаков листа (за исключением положения максимальной ширины и диагонали) и перекодированных признаков (максимальная корреляция - у отношения диагоналей) с типом волосков. Это может значить, что разделение видов по типу волосков не будет соответствовать возможным классификациям по другим признакам.
Таблица 4. Корреляции для измеренных признаков.
| BLANK.N | PLOSCH.N | TIP.POB | POBEG.N | N.L.SER | L.MEZHD | L.CHER | W.LOC | DIAG | L.MAX | W.MAX | PARTS | WHO | SMELL | TRICH | |
| BLANK.N | 1,00 | ,98 | ,05 | ,13 | -,11 | ,58 | ,61 | ,77 | ,67 | ,57 | ,43 | -,70 | -,74 | ,45 | ,64 |
| PLOSCH.N | ,98 | 1,00 | ,01 | ,24 | -,13 | ,50 | ,54 | ,73 | ,59 | ,46 | ,37 | -,62 | -,68 | ,36 | ,60 |
| TIP.POB | ,05 | ,01 | 1,00 | -,86 | -,18 | ,05 | ,03 | ,01 | ,01 | ,12 | -,03 | -,04 | -,28 | ,55 | -,36 |
| POBEG.N | ,13 | ,24 | -,86 | 1,00 | ,13 | -,15 | -,11 | -,03 | -,08 | -,27 | -,10 | ,06 | ,34 | -,68 | ,36 |
| N.L.SER | -,11 | -,13 | -,18 | ,13 | 1,00 | -,22 | -,30 | -,33 | -,19 | -,23 | -,24 | -,01 | ,22 | -,23 | ,03 |
| L.MEZHD | ,58 | ,50 | ,05 | -,15 | -,22 | 1,00 | ,55 | ,61 | ,54 | ,56 | ,37 | -,62 | -,62 | ,50 | ,42 |
| L.CHER | ,61 | ,54 | ,03 | -,11 | -,30 | ,55 | 1,00 | ,76 | ,86 | ,88 | ,84 | -,51 | -,66 | ,53 | ,39 |
| W.LOC | ,77 | ,73 | ,01 | -,03 | -,33 | ,61 | ,76 | 1,00 | ,81 | ,81 | ,72 | -,57 | -,79 | ,60 | ,53 |
| DIAG | ,67 | ,59 | ,01 | -,08 | -,19 | ,54 | ,86 | ,81 | 1,00 | ,90 | ,88 | -,51 | -,70 | ,55 | ,48 |
| L.MAX | ,57 | ,46 | ,12 | -,27 | -,23 | ,56 | ,88 | ,81 | ,90 | 1,00 | ,88 | -,52 | -,72 | ,64 | ,42 |
| W.MAX | ,43 | ,37 | -,03 | -,10 | -,24 | ,37 | ,84 | ,72 | ,88 | ,88 | 1,00 | -,27 | -,56 | ,47 | ,28 |
| PARTS | -,70 | -,62 | -,04 | ,06 | -,01 | -,62 | -,51 | -,57 | -,51 | -,52 | -,27 | 1,00 | ,60 | -,43 | -,54 |
| WHO | -,74 | -,68 | -,28 | ,34 | ,22 | -,62 | -,66 | -,79 | -,70 | -,72 | -,56 | ,60 | 1,00 | -,91 | -,43 |
| SMELL | ,45 | ,36 | ,55 | -,68 | -,23 | ,50 | ,53 | ,60 | ,55 | ,64 | ,47 | -,43 | -,91 | 1,00 | ,15 |
| TRICH | ,64 | ,60 | -,36 | ,36 | ,03 | ,42 | ,39 | ,53 | ,48 | ,42 | ,28 | -,54 | -,43 | ,15 | 1,00 |
Таблица 5. Корреляции для перекодированных признаков.
| WHO | PLOSCH.N | TIP.POB | POBEG.N | N.L.SER | L.MAX | PARTS | SMELL | TRICH | L.CH | L.Q | L.VY | L.W | |
| WHO | 1 | -0.68 | -0.28 | 0.34 | 0.22 | -0.72 | 0.6 | -0.91 | -0.43 | 0.17 | -0.63 | -0.47 | -0.48 |
| PLOSCH.N | -0.68 | 1 | 0.01 | 0.24 | -0.13 | 0.46 | -0.62 | 0.36 | 0.6 | -0.3 | 0.66 | 0.5 | 0.19 |
| TIP.POB | -0.28 | 0.01 | 1 | -0.86 | -0.18 | 0.12 | -0.04 | 0.55 | -0.36 | 0.14 | 0.13 | 0.17 | 0.26 |
| POBEG.N | 0.34 | 0.24 | -0.86 | 1 | 0.13 | -0.27 | 0.06 | -0.68 | 0.36 | -0.19 | 0 | -0.09 | -0.41 |
| N.L.SER | 0.22 | -0.13 | -0.18 | 0.13 | 1 | -0.23 | -0.01 | -0.23 | 0.03 | 0.31 | -0.42 | -0.01 | -0.04 |
| L.MAX | -0.72 | 0.46 | 0.12 | -0.27 | -0.23 | 1 | -0.52 | 0.64 | 0.42 | -0.24 | 0.31 | 0.26 | 0.46 |
| PARTS | 0.6 | -0.62 | -0.04 | 0.06 | -0.01 | -0.52 | 1 | -0.43 | -0.54 | 0.11 | -0.4 | -0.49 | -0.53 |
| SMELL | -0.91 | 0.36 | 0.55 | -0.68 | -0.23 | 0.64 | -0.43 | 1 | 0.15 | -0.04 | 0.47 | 0.36 | 0.52 |
| TRICH | -0.43 | 0.6 | -0.36 | 0.36 | 0.03 | 0.42 | -0.54 | 0.15 | 1 | 0.03 | 0.31 | 0.39 | 0.3 |
| L.CH | 0.17 | -0.3 | 0.14 | -0.19 | 0.31 | -0.24 | 0.11 | -0.04 | 0.03 | 1 | -0.2 | 0 | 0.15 |
| L.Q | -0.63 | 0.66 | 0.13 | 0 | -0.42 | 0.31 | -0.4 | 0.47 | 0.31 | -0.2 | 1 | 0.26 | 0.13 |
| L.VY | -0.47 | 0.5 | 0.17 | -0.09 | -0.01 | 0.26 | -0.49 | 0.36 | 0.39 | 0 | 0.26 | 1 | 0.52 |
| L.W | -0.48 | 0.19 | 0.26 | -0.41 | -0.04 | 0.46 | -0.53 | 0.52 | 0.3 | 0.15 | 0.13 | 0.52 | 1 |
Рис. 1. Корреляции для некоторых измеренных признаков.
Анализ распределения признаков (гистограммы) показывает, что большинство метрических признаков, скорее всего, не имеет нормального распределения (наблюдаются двувершинные гистограммы, см., например, рис. 2).
Рис. 2. Гистограмма распределения значений диагонали листа (DIAG)
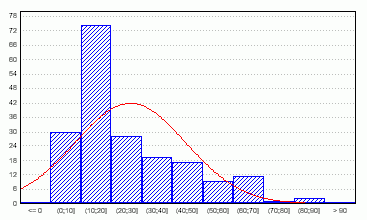
Дискриминантный анализ был проведен с целью выяснить, насколько хорошо классификация по признакам (1) количества лопастей; (2) запаха; (3) типа волосков соответствует классификациям, полученным по остальным признакам. Общие проценты корректности классификаций составляют 66,7%, 85,4%, 77,7% соответственно, что говорит о низком соотвествии классификаций (оптимальный процент корректности - от 90%). Распределение данных на плоскости дискриминантных корней для классификации по количеству лопастей позволяет увидеть некоторые группы, хотя весьма нечетко (рис. 3). Это - свидетельство более высокого соответствия признака количества частей остальным измеренным признакам.
Рис. 3. Распределение данных на плоскости корней дискриминантного анализа по количеству лопастей.
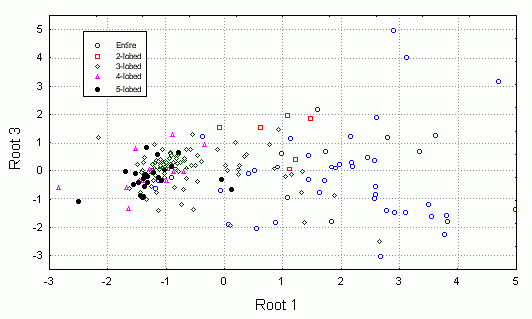
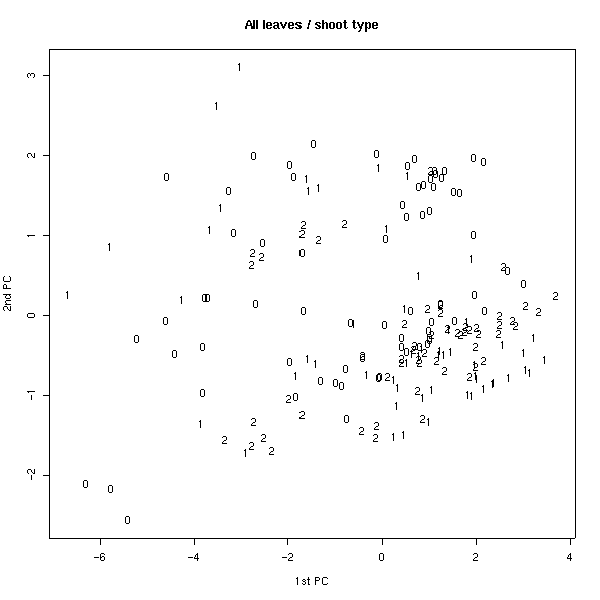
Был проведен также анализ главных компонент на основе измеренных признаков. На плоскости двух первых компонент удалось заметить, что по типам волосков листья образуют две четкие группы (рис 4). Для генеративных листьев эти группы менее четкие. Если провести многомерное шкалирование (вид кластерного анализа), то таких групп вовсе не наблюдается (рис. 5), что свидетельствует о неравном вкладе признаков в ╢правильную∙ классификацию - ведь кластерный анализ делается в предположении о равном весе всех признаков. По запаху также образуются группы, но значительно менее четкие (рис. 6). По участкам группы образуются плохо (рис. 7), по частям листа (рис. 8) - хорошо, а по типам побегов (рис. 9) не образуются. Такие результаты говорят о преобладающей роли признака типа волосков (TRICH) для классификации, хотя (см. рис. 6) группы разделены косым хиатусом и имеют нарастающую к их центрам плотность.
Рис. 4. Распределение по типам волосков (анализ главных компонент).
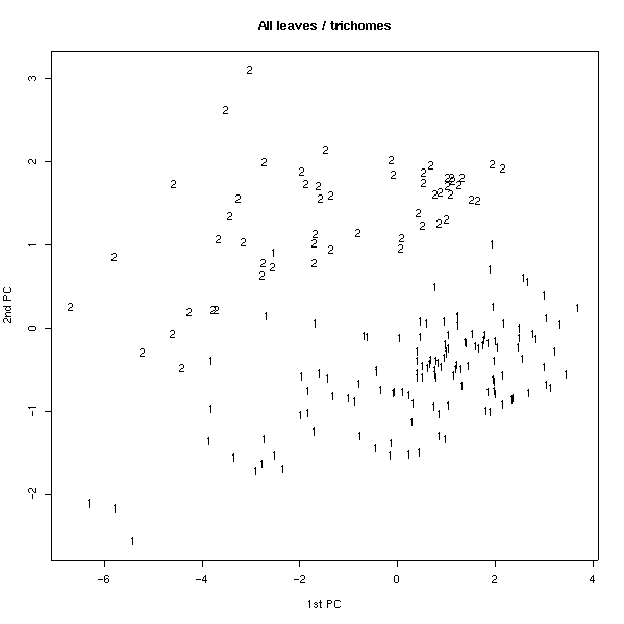
Рис. 5. Распределение по типам волосков (многомерное шкалирование).
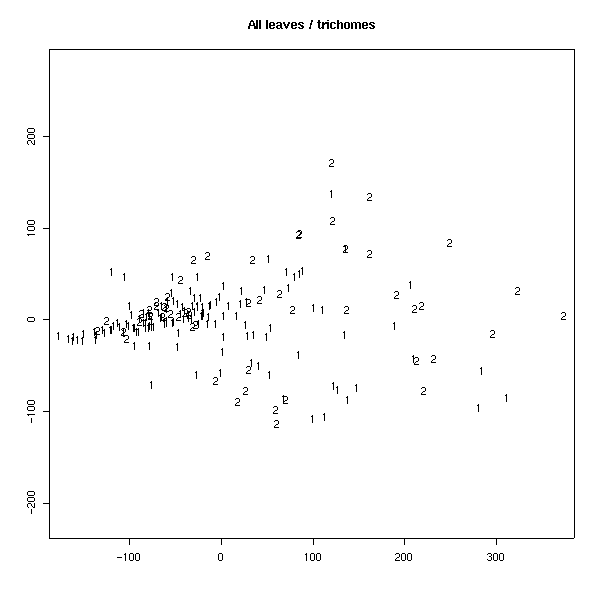
Рис. 6. Распределение по запаху (анализ главных компонент).
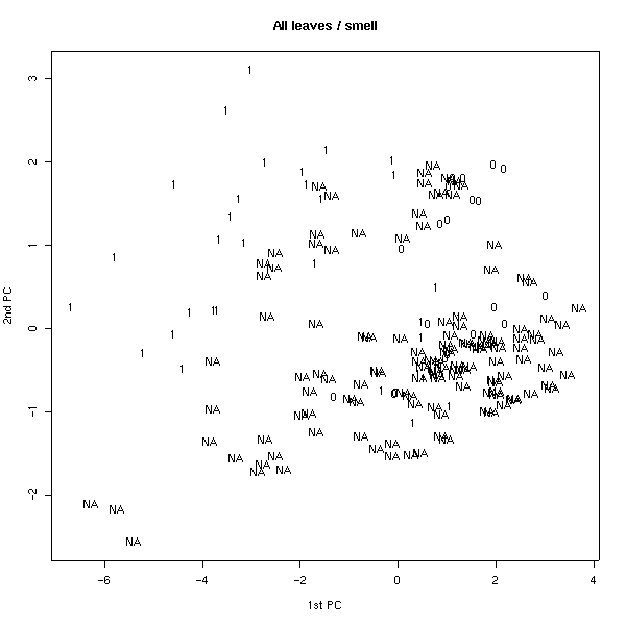
Рис. 7. Распределение по участкам (анализ главных компонент).
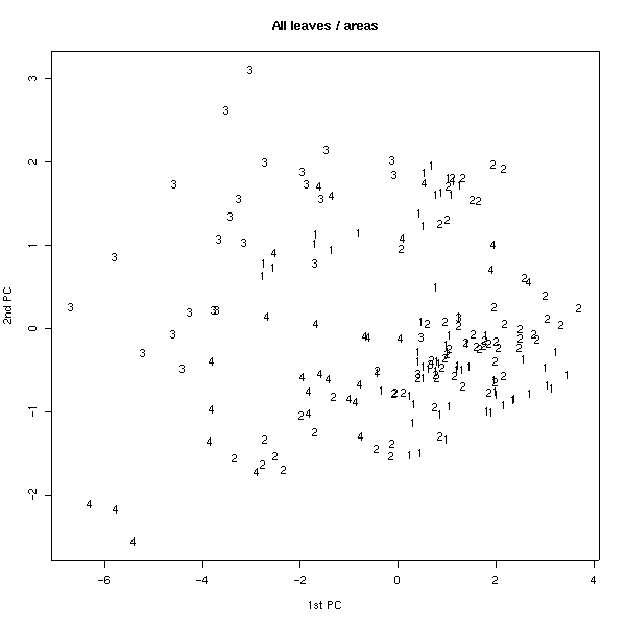
Рис. 8. Распределение по количеству частей листа (анализ главных компонент).

Рис. 9. Распределение по типам побегов (анализ главных компонент).
Наконец, факторные нагрузки (табл. 6, верхняя часть) говорят о том, что для данной классификации наиболее существенны 3 группы признаков: (1) количество лопастей + номер листа в серии; (2) тип волосков и (3) все (или любой из) метрические признаки листа.
Таблица 6. Факторные нагрузки на признаки (анализ главных компонент).
| Признаки/компоненты | Comp.1 | Comp.2 |
|
Анализ с участием признака ╢тип волосков∙ |
||
| номер листа в серии | 0.2416222 | -0.05802656 |
| длина междоузлия | -0.2486290 | 0.31864212 |
| длина черешка | -0.3773641 | -0.01087832 |
| положение наибольшей ширины | -0.3705915 | -0.12815823 |
| диагональ | -0.4192672 | -0.15282061 |
| длина листа | -0.4196109 | -0.19037944 |
| ширина листа | -0.3651233 | -0.35325888 |
| количество лопастей листа | 0.2935642 | -0.35206230 |
| тип волосков | -0.1694326 | 0.75512851 |
|
Анализ без участия признака ╢тип волосков∙ |
||
| номер листа в серии | 0.2423155 | 0.24658986 |
| длина междоузлия | -0.2471571 | 0.81583012 |
| длина черешка | -0.3804359 | 0.30443867 |
| положение наибольшей ширины | -0.3768565 | -0.30598545 |
| диагональ | -0.4272458 | -0.20585166 |
| длина листа | -0.4284686 | -0.16171701 |
| ширина листа | -0.3784350 | 0.10588842 |
| количество лопастей листа | 0.2900120 | 0.08697706 |
Для того, чтобы проверить гипотезу о ведущей роли признаков волосков, мы устранили этот признак из первоначальных данных. Анализ главных компонент в этом случае дал гораздо менее четкие результаты (рис. 10, 11); кроме того, возросла факторная нагрузка на признак длины междоузлий (табл. 6, нижняя часть).
Рис. 10. Распределение по типам волосков (анализ главных компонент, без TRICH).
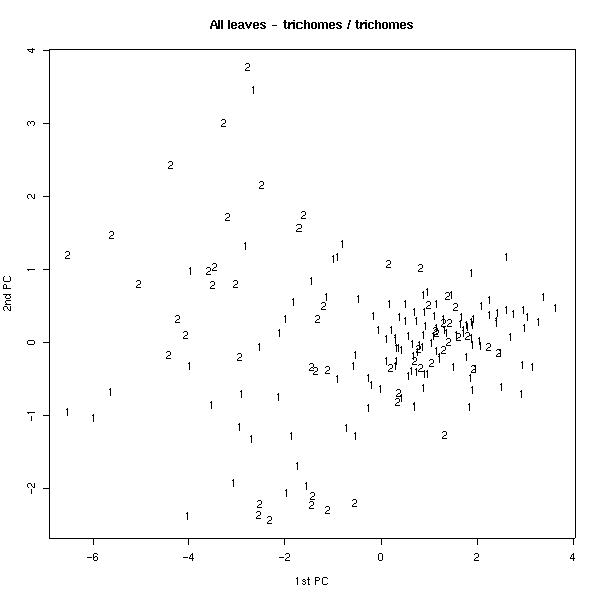
Рис. 11. Распределение по участкам (анализ главных компонент, без TRICH).
Попытки классификации, основанные на выделении максимальных листьев в каждой серии, результатов не дали.
Анализ главных компонент был проведен также для перекодированных признаков. Как показывает таблица факторных нагрузок для этого случая (табл. 7), нагрузки распределились более или менее равномерно между перекодированными признаками (наибольшая - у отношения диагоналей) и номером листа в серии. Тип волосков существенной нагрузки не несет, что может говорить об относительно большей классифицирующей способности перекодированных признаков. Большая нагрузка на номер листа в серии может говорить за то, что в данном случае разделение листьев на группы зависит от положения листа внутри серии. Однако группы не образуются (рис. 12-14). Это говорит, скорее всего, за то, что (а) классификация с использованием только перекодированных признаков не соответсвует разделению видов по типу волосков, либо за то, что (б) измеренные серии образуют не две (по типам волосков), а большее число групп (менее вероятно, поскольку классификация здесь зависит от положения листа в серии - см. выше).
Таблица 7. Факторные нагрузки на признаки (анализ главных компонент, перекодированные признаки).
| Признаки/компоненты | Comp.1 | Comp.2 |
| номер листа в серии | -0.22318424 | 0.59220331 |
| количество частей | -0.45150048 | 0.07837735 |
| тип волосков | 0.31423412 | -0.30385218 |
| приподнятость листа | 0.26016841 | 0.62507308 |
| вытянутость листа | 0.04268869 | -0.35169929 |
| отношение диагоналей | 0.54986530 | 0.02993812 |
| яйцевидность листа | 0.52510119 | 0.18847026 |
Рис. 12. Распределение по типам волосков (анализ главных компонент, перекодированные признаки).
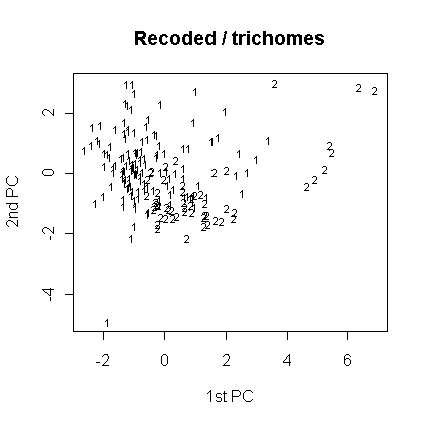
Рис. 13. Распределение по участкам (анализ главных компонент, перекодированные признаки).
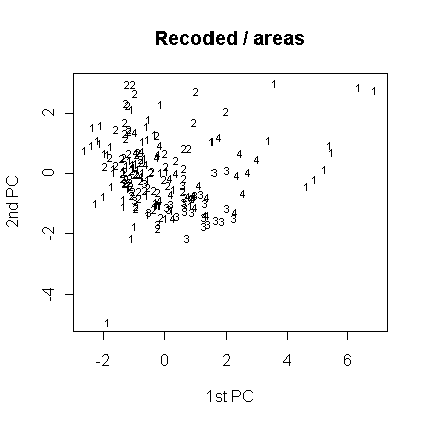
Рис. 14. Распределение по количеству частей листа (анализ главных компонент, перекодированные признаки).
1) Метрические признаки листа признаки очень изменчивы, слабо коррелируют с известными диагностическими признаками и поэтому плохо подходят для классификации. Наиболее перспективными для классификации являются положение максимальной ширины и диагональ, а также длина междоузлия, а из перекодированных - отношение теоретической и измеренной диагонали.
2) Из неметрических признаков наиболее перспективен признак количества лопастей листа.
3) Классификация по волоскам наиболее адекватна представлениям, сложившимся в литературе, но не совпадает с классификацией, полученной с использованием перекодированных признаков. Последняя зависит также и от положения листа в серии, что говорит за то, что эта классификация не имеет таксономического значения.
4) Классификация по запаху отличается сильной субъективностью и нуждается в дальнейшей проверке.
5) Площадки различаются слабо, что может говорить либо об отсутствии двух видов, либо об их сосуществовании (и, возможно, гибридизации) на одной площадки, либо о еще более сложной таксономической структуре.
6) Типы побегов не различаются при классификации.
7) В дальнейшем следует: уменьшить количество измеряемых метрических признаков, разработать методику для более объективного определения запаха и новые методы перекодирования признаков. Следует также увеличить число наблюдений за счет большего количества точек (площадок).
Авторы работы благодарят С. Глаголева и А. Квашенко за помощь в проведении работы; А. Багдалову, Н. Кулыгину, И. Юфрякова, А. Акайкина, Н. Станцо, С. Лосек за проведенные в природе измерения и А. Оскольского (БИН РАН) за ценные замечания, высказанные в процессе проведения работы, а также помощь в предоставлении литературы.
А. С. Зернов. Определитель растений Северо-Западного Закавказья. М., в печати.
Н. П. Кренке. Феногенетическая изменчивость // Тр. Биол. ин-та им. К. А. Тимирязева. Т. 1. М., 1933. 860 с.
J. Ackerfield. Trichome morphology in Hedera (Araliaceae) // Edinb. J. Bot. 2001. P. 259-267.
R. Ihaka, R. Gentleman. R: A Language for Data Analysis and Graphics // J. Comp. Graph. Stat. 1996 Vol. 5. N 3. P. 299-314.
STATISTICA for Windows // Computer program manual. StatSoft, Inc. 1999. Tulsa, OK.
P. Vargas, H. A. McAllister, C. Morton, S. L. Jury, M. Wilkinson. Polyploid speciation in Hedera (Araliaceae): phylogenetic and biogeographic insights based on chromosome counts and ITS sequences // Pl. Syst. Evol. 1999. Vol. 219. P. 165-179.
Webmaster: Полина Волкова