Drosera - один из самых крупных по количеству и разнообразию жизненных форм родов насекомоядных растений. В нем известно более 90 видов. Центр разнообразия рода Drosera находится в Австралии и Новой Зеландии (Crowder et al., 1990). В нашей стране обитает 4 представителя этого рода - D. rotundifolia L., D. anglica Huds., D. х obovata Mert. et Koch, D. intermedia Hayne.
Целью нашей работы являлось изучение движений листьев трёх видов: Drosera rotundifolia L., D. anglica Huds. и D. obovata Mert. et Koch.
Для того, чтобы оценить, существует ли внутригрупповая синхронизация ритма движений листьев, полученные числовые значения усреднялись для всех растений в определённый момент времени. Этот показатель был назван “усреднённым положением листа”. На каждой площадке наблюдения проводились 3-4 суток. Этот срок определялся началом пересыхания растений из-за притаптывания прилегающих сфагновых мхов. Лист, регулярно менявший своё положение, считался движущимся. Неподвижными считались листья, не менявшие своё положение ни разу в ходе эксперимента. Не учитывались : а) молодые листья, б) старые листья, в) листья, поймавшие насекомое и начавшие сгибаться в ответ на раздражение листовой пластины. Лист считался захватившим насекомое в случае, если насекомое находилось на листе более 8 часов. В случае, если насекомое за это время освобождалось, движения листа продолжали учитываться за исключением того периода, когда на листе находилось насекомое. Если молодой лист в процессе наблюдения успевал развиться полностью и начинал ритмические движения, он наблюдался как движущийся лист.
Для каждого наблюдаемого растения были описаны условия его произрастания. Учитывалась освещённость местообитаний, социальная структура, жизненность растения, удалённость от грунтовых вод, ход температуры. Под жизненностью подразумевалась интенсивность окраски растения и наличие на нём цветоноса. Под социальной структурой подразумевалось положение растения относительно других : а) одиночное, б) произрастает в тесной группе. Относительная удалённость от грунтовых вод учитывалась только на Беломорском побережье губы Чупа, поскольку на Сокольском болоте Вологодской области наблюдаемый участок имел сглаженный микрорельеф. Средний прирост растений вычислялся из данных, полученных при измерения прироста за всё время жизни модельных растений, взятых на 20 площадках 1х1 м.
Для обработки результатов использовался метод хи-квадрат, реализованный в программе Statistica 5.0 (Statsoft Inc., 1984-1995).
В ходе исследования также была проведена морфометрия растений D. rotundifolia на Сокольском болоте в Вологодской области. Измерялись диаметры розеток растений и листьев, количество растений в вегетативном и в цветущем состоянии на площадках размером 1х1 м. Таким образом было исследовано 25 площадок. На каждой площадке учитывалась освещённость и окраска каждого растения по 3-балльной шкале (табл.1).
Таблица 1.
| 1 | 2 | 3 | |
| освещённость | наиболее освещённые места | постоянно слабо затенённые или затенённые частично в течение суток места | места с постоянным сильным затенением |
| интенсивность окраски растения | листья окрашены в интенсивно красный цвет | листья с нормальной окраской, при которой желёзки листа окрашены в красный цвет, а пластина - в зелёный | листья с зелёной окраской, у которых практически отсутствовала красная окраска желёзок |
На основе полученных данных по движениям листьев, находившихся в разных условиях, были построены графики, отражающие временную зависимость количественного показателя положения листьев (усреднённое по всем листьям). На графиках следует учитывать, что положение неподвижных листьев было примерно равно единице. Движущиеся листья колебались от положения 1 до положения 2. Поэтому в нижней точке графики движущихся и недвижущихся листьев часто совпадают.
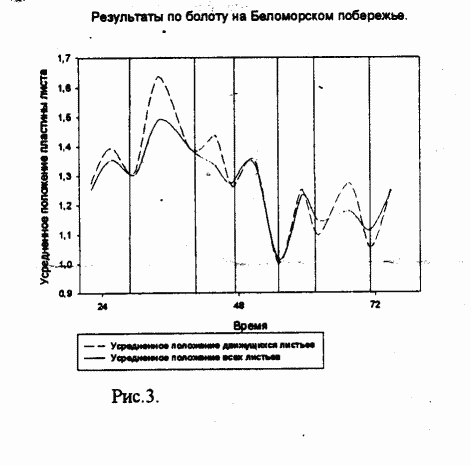
В ходе работы было установлено, что изученные виды Drosera имеют движения листьев, не связанные с улавливанием насекомых, причём такие движения ритмичны. На Беломорском побережье период движения листьев был примерно равен 15 часам (рис.1), причём он сильно варьировал, а в Вологодской области - 12 часам (рис.2).
Рисунок 1.
Сплошной линией обозначено усреднённое положение движущихся листьев; пунктирной линией обозначено положение всей листьев.
Рисунок 2.
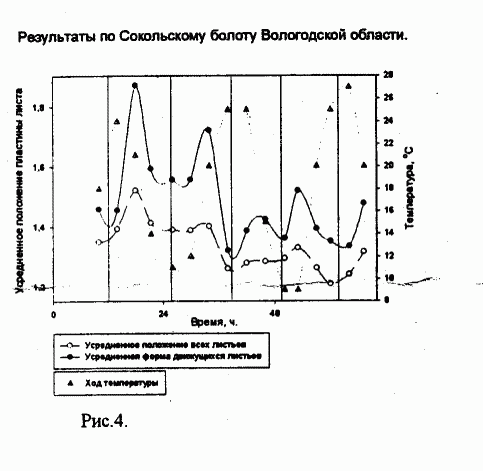
Обозначения совпадают с предыдущим рисунком; точечной линией обозначен ход температуры.
Эти различия, как нам представляется, могут быть связаны с разным фотопериодом в этих районах. Для объяснения полученных результатов нами было выдвинуто несколько рабочих гипотез:
В работе на побережье губы Чупа проведено сравнение движения листьев видов Drosera rotundifolia, D. anglica и D. obovata. В Вологодской области такого сравнения не проводилось, т.к. D. anglica и D. obovata отсутствовали.
В таблице 2 приведены данные по движениям листьев разных видов.
Таблица 2. Количество движущихся и наподвижных листьев, а также пойманных
насекомых у разных видов Drosera.
| общее кол-во листьев | кол-во насекомых | кол-во движущихся листьев | Кол-во недвигающихся листьев | кол-во старых, молодых и пр. неучтенных листьев | |
| Drosera rotundifolia | |||||
| одиночных | 37 | 2 | 5 | 15 | 15 |
| групповых | 16 | 9 | 2 | 1 | 4 |
| Всего | 53 | 11 | 7 | 16 | 19 |
| D. anglica | |||||
| одиночных | 35 | 18 | 5 | 8 | 4 |
| групповых | 42 | 23 | 4 | 2 | 13 |
| Всего | 77 | 41 | 9 | 10 | 17 |
| D. obovata | |||||
| одиночных | 56 | 13 | 2 | 5 | 36 |
| групповых | 21 | 7 | 1 | 5 | 8 |
| Всего | 77 | 20 | 3 | 10 | 44 |
Из таблицы видно, что у одиночных растений всех трёх видов общее количество листьев значительно больше, чем у растущих в группе. Также видно, что D. anglica более успешна в ловле насекомых, чем другие виды, при этом количество движущихся листьев у разных видов сильно не различается.
С помощью критерия хи-квадрат была изучена взаимосвязь количества движущихся листьев и различных факторов, - освещённости, социальной структуры, жизненности растения, удалённости от грунтовых вод. Достоверная взаимосвязь получена только для удалённости от грунтовых вод (с уровнем достоверности 0,028) и для освещённости (с уровнем достоверности 0,022), что может быть связано с тем, что в период наблюдений стояла сухая погода, и количество влаги было связано со скоростью высыхания. Полученные данные сочетаются с результатами изучения движений листьев Abutilon, который также увеличивает интенсивность движений при снижение поступления воды (Koide et al., 1994).
C помощью метода хи-квадрат проводилось также исследование взаимосвязи разных факторов. Рабочая гипотеза о том, что чем более освещено растение, там интенсивнее его окраска, не подтвердилось. Наблюдения показали, что окраска растения зависит скорее от того, насколько большую роль в обеспечение растения ресурсами играет насекомоядность. При этом, по видимому, наиболее благоприятна окраска пластины листа в зелёный цвет, а желёзок - в красный. Полностью красные листья были либо стареющими, либо находились в неблагоприятных условиях, а зелёные - либо молодыми, недавно развернувшимися, либо также находились в неблагоприятных условиях.
В ходе наблюдений замечено влияние листа, сгибающегося вследствие поимки насекомого, на лист, не связанный с ним. При этом листья должны располагаться в непосредственной близости друг к другу, но не обязательно, чтобы они принадлежали одному растению. Лист, не поймавший насекомое, но расположенный близко к листу, завладевшему добычей, поднимает желёзки и начинает сгибать пластину листа, т.е. приходит в положение “2”. Это явление может быть объяснено, например, выделением сгибающимся листом летучих веществ.
Брюс, В.Г. Влияние периодических внешних воздействий. - “Биологические часы” под ред. С.Э. Шноля, М., Мир, 1964.
Bunning, E. Untersuchungen uber die autonomen tagesperiodischen Bewegungen der Primarblatten von Phaseolus multiflorus Jahrb. //Wiss.Bot.. 1931. Vol. 75. P. 439-480.
Crowder, A.A., Pearson, M.C., Grubl, P.J., Landlois, P.H. Biological flora of the British Isles. //Journal of ecology. 1990. Vol. 78. P. 233-267.
Highkin, H.R. Temperature induced variability in peas. //Am. J. Bot.. 1958. Vol. 45. P. 628-631.
Hillman, W.S. Injury of tomato plants by continuous light and unflorable photoperiodic cycles. //Am.J Bot.. 1956. Vol. 43. P. 89-96.
Juniper, B.E., Robins, R.J., Joel, D.M. The Carnivorous Plants. L., Academic Press. 1989.
Koide, R.T., Schreiner, R.P. Alteration of leaf movement
of Abutilon theophrasti (Malvaceae) by mycorrhizal infection. //Functional
ecology. 1994. Vol. 8. N.3. P. 384-388.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |