Род Alnus Mill. включает 30–40 видов, распространенных в умеренных широтах Северного полушария (Комаров, 1936). Некоторые виды заходят также в Южную Америку до Аргентины и Чили, а в Азии доходят до Бенгалии и Северного Вьетнама, но встречаются там только в горах. На севере отдельные виды доходят до лесотундры и тундры, а в горах поднимаются до субальпийского пояса.
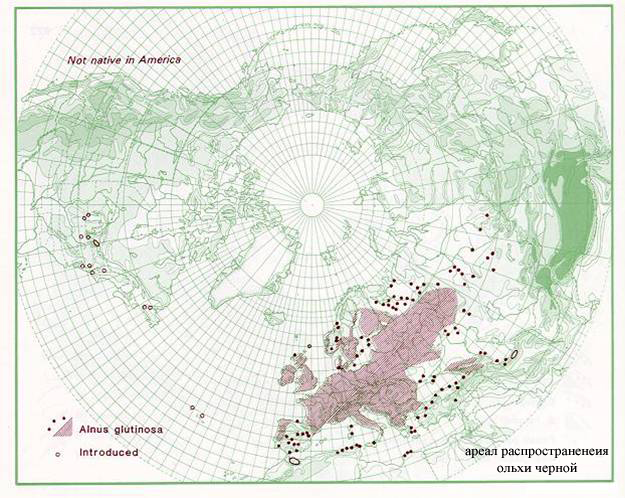
Большинством систематиков (см., например, Черепанов, 1955) род Alnus разделяется на несколько секций: (1) Гимнотирзус (Hymnothyrsus) - самая крупная секция, к которой принадлежит, например, Alnus glutinosa. Представители секции (2) Гаплостахис (Haplostachys) произрастают только вблизи морских побережий. Секция (3) Проскеймостемон (Proskeimostemon) распространена преимущественно в Европе (рис. 1, 2) и включает Alnus incana и A. kolaensis (Орлова, 1954; Цвелев, 2002). Следует сказать, однако, что данные анализа ДНК (Navarro et al., 2003) противоречат вышеуказанной системе.
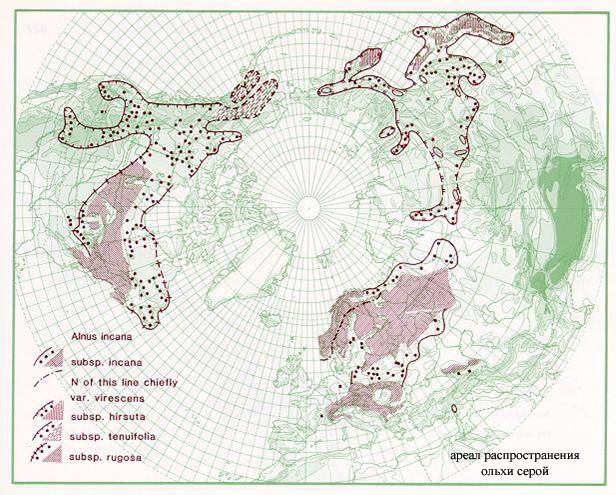
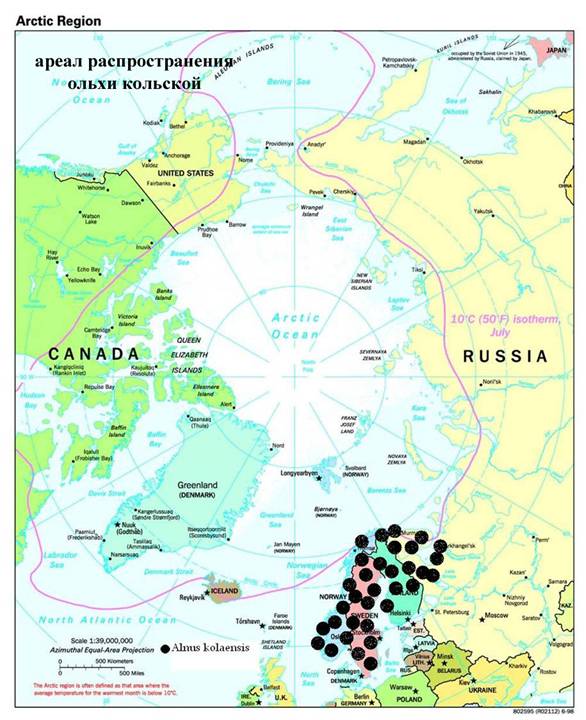
Alnus incana – самый распространенный вид ольхи Евразии. Широко известен ее полиморфизм. Существуют, к примеру, формы, близкие к A. glutinosa. Одной из таких форм является кольская ольха, часто рассматривающаяся как особый вид Alnus kolaensis Orlova. Эта ольха произрастает на юго-востоке Фенноскандии и Норвегии, на Кольском полуострове и на Карельском побережье Белого моря (рис. 3). Различные систематики по-разному понимают статус кольской ольхи. Так, Н.И. Орлова (1954) считает ее отдельным видом, а O. Nilsson (2000) и А.К. Скворцов (1959) считают Alnus kolaensis подвидом A. incana. Кроме того, некоторые авторы указывают на гибридное происхождение A. kolaensis (которую в этом случае рассматривают как гибридный вид). А. Кундзиньш (1969) и Н.Н. Цвелев (2002), например, упоминают в этом контексте западноевропейский природный гибрид Alnus x hybrida (A. glutinosa x A. incana). Н.Н. Цвелев (2002) считает, что и A. kolaensis – продукт гибридизации этих же двух видов. С другой стороны, O. Nilsson (2000) рассматривает A. x hybrida и A. kolaensis как отдельные формы. Дело осложняется тем, что в настоящее время на большей части ареала ольхи кольской не встречается один из предполагаемых "родителей" – A. glutinosa. Наконец, Е.В. Банаев и М.А. Шемберг (2000) считают, что некоторые сибирские экземпляры A. hirsuta неотличимы от A. kolaensis.
Целью данной работы является выяснение таксономической принадлежности беломорских популяций ольхи.
Существует множество способов анализа: хромосомный, морфологический (Банаев, Шемберг, 2000), анализ ДНК (King et al., 1998; Navarro et al., 2003), а также достаточно новый метод анализа форм, появившийся в девяностых годах прошлого века – геометрическая морфометрия (Павлинов, Микешина, 2002). Подавляющее большинство публикаций, связанных, так или иначе, с методами геометрической морфометрии, выходит на английском языке, причем применению этого подхода для анализа формы ботанических объектов посвящено лишь несколько статей. В книге «Ольха в Сибири и на Дальнем Востоке России» (2003) авторы применяли методы классической морфометрии для сравнения формы листьев различных видов ольхи. Из 17 исследованных характеристик 11 являлись характеристиками листовой пластинки. По мнению авторов, «...лист является одним из главных органов, связанных с функционированием растений, поэтому его признаки должны подвергаться значительному давлению отбора...». Поэтому мы решили провести анализ двумя методами, решая при этом сразу две задачи: (1) – выяснение взаимоотношения Alnus incana, Alnus glutinosa и Alnus kolaensis, а также (2) – сравнение результатов, полученных методами классической и геометрической морфометрии.
Сбор материала проводили в июне-октябре 2002 и 2003 годов в Удомельском и Вышневолоцком районах Тверской области Лоухском районе республики Карелия. Для получения контуров листовой пластинки с каждого дерева брался максимальный лист листовой серии удлиненного побега и обводился на листе бумаги. Полученные контуры листьев вводили в компьютер через планшетный сканер. Для всех исследованных растений были также сделаны промеры длины и ширины листовой пластинки, расстояния от ее основания до положения ее наибольшей ширины количество пар жилок первого порядка с левой стороны, а так же оценивалось опушение листа и побега по трехбалльной шкале. Для этих промеров мы выбирали листья, которые были использованных для получения контура. Таким образом, данные для классической и геометрической морфометрии собирались для каждого конкретного листа. Для характеристики формы листа использованы 8 меток, расположенных на его контуре (рис. 4).
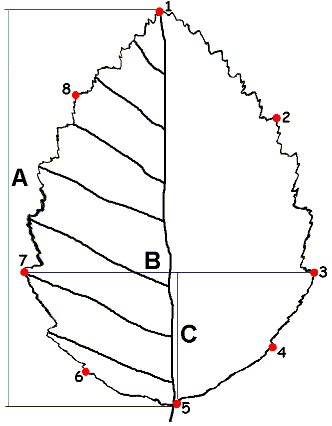
A -- длина листовой пластинки, B -- ширина листовой пластинки, C -- "положение наибольшей ширины"; пронумерованные точки соответствуют расположению меток для геометрической морфометрии
В нашем случае для фиксации меток использовались не только геометрические, но и биологические соображения: метки расставлялись в точках наибольшей кривизны контура. Использование таких меток позволяет сравнивать форму объектов, получая представления о биологическом смысле обнаруженных различий. Координаты точек снимали с изображений и записывали в файл данных при помощи экранного дигитайзера tpsDig. Координаты эталонной конфигурации, а также значения главных, относительных и частных трансформаций, характеризующие меру отличия образца от эталона, вычисляли при помощи программы tpsRelw. Исходные координаты экземпляров нормировали относительно эталонной конфигурации при помощи прокрустова наложения. Усредненные контуры листовой пластинки для каждого вида получали путем суперимпозиции при помощи программы tpsSuper. Редактирование и конвертирование файлов данных мы производили при помощи вспомогательной программы tpsUtil.
В связи с тем, что у нас было много переменных мы также применили многомерное шкалирование, анализ главных компонент матриц значений частных деформаций, шкалированных результатов морфологических промеров и кластерный анализ были проделаны при помощи пакета программ R 1.8.1.
В пространстве двух измерений на графике, сделанном по выводам геометрической морфометрии, образуется три облака (рис. 5)). Два из них представляют собой виды A. glutinosa (G) и A. cordata (C). Как видно на графике, есть некоторое количество «выбросов», которые, возможно, объясняются погрешностями в измерениях. Оставшееся облако представляет собой особи видов A. incana из Средней России и беломорские образцы, которые перекрываются значительно сильнее между собой, чем с «облаками» других видов.

На графике, полученном методами классической морфометрии, виды образуют отдельные «облака», которые в той или иной степени перекрываются (рис. 6).
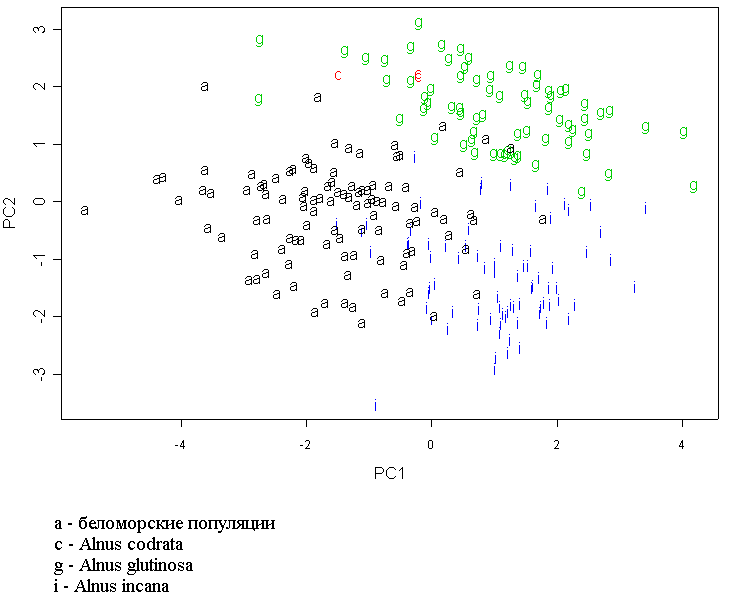
При классификации отдельных особей методами геометрической морфометрии мы получили неожиданные результаты. Возможно, такое разделение связано с наличием выемки на листьях Alnus glutinosa. Однако вопрос о таксономической принадлежности беломорских популяций остается открытым. Скорее всего, мы имеем дело с одной из форм Alnus incana (например, Alnus incana var. argentata).
Сбор большей части данных для настоящей работы проводился в рамках летних биологических практик Московской Гимназии на Юго-Западе N 1543. Мы благодарим руководителей практик С.М. Глаголева, А.Н. Квашенко, а также участников практик И.С. Юфрякова, Е.А. Кост и В.С. Рудакову, активно помогавших нам в полевой работе.
Банаев Е.В., Шемберг М.А. Ольха в Сибири и на Дальнем Востоке России // Новосибирск: изд-во Сибирского отделения РАН, 2000. 100 с.
Давидов М.В. Лесоводственно-биологическая характеристика ольхи // М.: Лесная промышленность, 1972. С. 4--13.
Комаров В.Л. Род Ольха – Alnus Gaertn. // Флора СССР. М.;Л.: Издательство АН СССР, 1936. С. 306--319.
Кундзиньш А. Исследование рода ольхи (Alnus Gaertn.) в Латвийской ССР // Автореф. дисс. ... доктора сельскохозяйственных наук. 1969. С. 28--39.
Орлова Н.И. Новый вид ольхи с Кольского полуострова // Бот. мат. Герб. БИН. 1954. Т. 16. С. 70--75.
Павлинов И.Я., Микешина Н.Г. Принципы и методы геометрической морфометрии // Журн. общ. биол. 2002. Т. 63. N 6. С. 473--493.
Раменская М.Л., Андреева В.Н. Alnus Mill. --- Ольха // Определитель высших растений Мурманской области и Карелии. Л.: Наука, 1982. С. 169--170.
Скворцов А.К. Новые флористические находки в районе Денежкина Камня (Северный Урал) // Бот. мат. Герб. БИН. М.;Л.: Изд-во АН СССР, 1959. Т. 19. С. 558--571.
Соколов Д.Д., Филин В.Р. Alnus Mill. --- Ольха // Определитель сосудистых растений окрестностей ББС МГУ. М.: Изд-во НЭВЦ ФИПТ, 1996. С. 76.
Цвелев Н.Н. О родах Betula L. и Alnus Mill. (Betulaceae) в Восточной Европе. // Новости систематики высших растений. Т. 34. Спб., 2002. C. 47-70.
Черепанов С.К. Система рода Alnus Mill. s. str. и близких к нему родов // Бот. мат. герб. БИН. М.;Л.: Изд-во АН СССР, 1955. Т. 17. С. 91--105.
AdamsC.D., Rohlf J.F., Slice D.E. Geometric morphometrics: ten years of progress following the "Revolution" // Italian Journal of Zoology. 2003.
Furlow J.J. The systematic of American species of Alnus (Betulaceae) // Rhodora. Vol. 81. N. 825. 1979. P. 1--69.
King A.R., Ferris C. Chloroplast DNA phylogeography of Alnus glutinosa (L.) Gaertn. // Molecular ecology. 1998. Vol. 7. P. 1151--1161.
Navarro E.J., Bousquet A., Moiroud A., Munive D., Piou Ph. Normand. Molecular phylogeny of Alnus (Betulaceae), inferred from nuclear ribosomal DNA ITS sequences // Plant and Soil. Vol. 245. P. 207--217.
Nilsson O. Alnus Mill. // Flora Nordica. Stokholm, 2000. Vol. 1. P. 203--208.
Walters S.M. Alnus Mill. // Flora Europea. 2nd ed. Cambridge, 1993. Vol. 1. P. 68--70.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |