Временные водоемы и, в частности, наскальные лужи - интересный объект для экологических исследований. Довольно простая структура сообществ и небольшое число видов позволяет подробно изучить факторы, влияющие на распределение видов в водоемах. Кроме того, лужи (как и острова, на которых они находятся) – идеальный объект для изучения влияния распространения и вымирания видов на состав сообществ.
Наиболее подробно фауна беспозвоночных и распределение видов в наскальных лужах морских побережий были исследованы на островах Балтийского моря и балтийских побережьях Финляндии и Швеции (Ranta, 1979, 1982; Ranta, Tjossem, 1987; Bengtsson, 1986, 1988).
В этих работах показано, что на распределение видов (в том числе, дафний D. magna, D. pulex, D. longispina) влияют такие факторы, как соленость и степень постоянства водоема (показатель, в свою очередь зависящий от глубины). Экологические ниши различных видов (в том числе близких) в отношении абиотических факторов различаются, однако обычно сильно перекрываются. В наиболее соленых лужах встречается D. magna. D. pulex чаще других видов встречается в лужах, пересыхающих летом.
Межвидовая конкуренция, видимо, не относится к числу важных факторов, влияющих на распределение видов. Так, в длительных опытах в искусственных лужах (контейнерах объемом 50 л) в природных условиях было показано, что в течение двух лет часто все три исследованных вида дафний продолжали сосуществовать /2,3/.
Некоторое влияние на распределение видов оказывает наличие хищников. Хищниками для кладоцер в изученных сообществах являются тритоны, девятииглая колюшка (зрительные позвоночные хищники), а также гладыши, кориксиды, жуки-плавунцы и другие беспозвоночные. Отмечалось, что D. longispina чаще встречается в лужах, где присутствуют хищники /3,7/.
Многолетние исследования на мелких водоемах проводятся редко, однако по наскальным лужам шведских островов есть данные четырехлетних наблюдений /3/. Существенная черта наскальных водоемов – высокие темпы вымирания популяций и темпы колонизации (около 10% на популяцию в год). В этих условиях сильное влияние на состав сообществ и генетическую структуру популяций должен оказывать эффект основателя, усиливающийся благодаря последующей локальной адаптации. Случайно проникнув в водоем, популяция может “монополизировать” его вследствие быстрой адаптации к данным условиям и накоплению банка покоящихся яиц, что может служить мощным барьером для проникновения конкурентов /4/.
Мы провели такие же работы на островах Белого моря. При этом мы ставили перед собой следующие задачи:
В 2002 г. проводилась подготовительная работа, были описаны 54 лужи на шести островах Керетского архипелага (о. "Скелетов", о. Виченная Луда, о. "Луда Ожидания", о. Седловатая Луда, о. Черемшиха, о. "Средняя Одинокая Луда"). На каждом острове описывалось от одной до 13 луж.
В 2003 г. работа была повторена. Описано 79 луж: 41 лужа на 7 островах Керетского архипелага (о. "Большая Одинокая Луда", о. "Средняя Одинокая Луда", о. "Черепаха", о. Сидоров, о. "Скелетов", о. Иваньков, о. "Гроздовник") и 38 луж на островах Кемь-Лудского архипелага (о. Коржничиха, о. "Кораблик", о. Большой Асафьев), на каждом острове описывалось от 4 до 31 лужи. К сожалению, посетить повторно удалось лишь четыре лужи, исследованные в 2002 г.
Описание лужи происходило следующим образом:
С использованием определителя пресноводных беспозвоночных России /1/ были определены виды ветвистоусых (Cladocera) и составлен видовой список с частотами встречаемости видов (табл. 1). Остальные группы беспозвоночных пока определены нами лишь до отрядов и до семейств (исключение составляют коловратка Hexarthra fennica, в массовом количестве найденная в одной из луж, и присутствовашие в нескольких лужах личинки комаров-хаоборусов, относящиеся к виду Chaoborus obscuripes). В 2003 г. до вида определены также жуки (табл. 2).
В ходе статистической обработки данных были проведены:
Видовой список ветвистоусых, встреченных нами в исследованных лужах в 2003 году, включает 10 видов из 5 семейств. Для каждого вида были определены частоты встречаемости (в %) в исследованных лужах (табл. 1).
| вид | частота встречаемости (%) | |||
| Керетский архипелаг | Кемьлудский архипелаг | |||
| 2002 год | 2003 год | 2003 год | 2002 год и 2003 год | |
| сем. Polyphemidae | ||||
|
Polyphemus pediculus |
5 | 7,3 | 18,4 | 10,2 |
| сем. Daphniidae | ||||
|
Daphnia magna |
43 | 41,4 | 34,2 | 39,5 |
|
Daphnia pulex |
13 | 19,5 | 5,2 | 12,5 |
|
Daphnia curvirostris |
17 | 0 | 10,5 | 4 |
|
Daphnia longispina |
10 | 9,7 | 7,8 | 9,1 |
|
Ceriodaphnia quadrangula |
38 | 19,5 | 10,5 | 22,6 |
|
Ceriodaphnia cf. dubia |
3,3 | --- | --- | 3,3 |
|
Scapholeberis mucronata |
20 | 19,5 | 15,7 | 18,4 |
| сeм. Chydoridae | ||||
|
Chydorus sphaericus |
38 | 60,9 | 50 | 49,6 |
|
Alonella excisa |
1,7 | --- | --- | 1,7 |
| сeм. Macrothricidae | ||||
|
Macrothrix hirsuticornis |
3,3 | 2,4 | 2,6 | 2,7 |
|
Streblocerus serricaudatus |
3,3 | --- | --- | 3,3 |
| сем. Bosminidae | ||||
|
Bosmina longispina s.l. |
9 | 14,6 | 13,1 | 12,2 |
| систематическая группа | частота встречаемости (%) | ||
| Керетский архипелаг | Кемьлудский архипелаг | общее | |
|
Coleoptera |
56 | 55,2 | 55,6 |
|
Ditiscidae |
51,2 | 55,2 | 53,2 |
|
Hygrotus impressopunctatus |
0 | 5,2 | 5,2 |
|
Hygrotus novemlineatus |
0 | 5,2 | 5,2 |
|
Hydroporus erythrocephalus |
4,8 | 2,6 | 3,7 |
|
Hydroporus geniculatus |
0 | 2,6 | 2,6 |
|
Hydroporus morio |
41,4 | 47,3 | 46,2 |
|
Hydroporus obscurus |
7,3 | 5,2 | 6,2 |
| Hydroporus striola | 4,8 | 7,8 | 12,6 |
|
Hydroporus tristis |
0 | 5,2 | 5,2 |
|
Hydroporus umbrosus |
2,4 | 0 | 2,4 |
|
Agabus thomsoni |
19,5 | 13,1 | 16,3 |
|
IIybius aenescens |
2,4 | 0 | 2,4 |
|
llybius vittiger |
0 | 2,6 | 2,6 |
|
Rhantus suturellus |
12,1 | 0 | 12,1 |
|
Gyrinus opacus |
2,4 | 0 | 2,4 |
|
Hydrophilidae |
24,3 | 28,9 | 26,6 |
|
Hydraenidae |
0 | 2,6 | 2,6 |
|
Hemiptera |
26,8 | 18,4 | 22,6 |
|
Corixidae |
21,9 | 18,4 | 20,1 |
|
Gerridae |
7,3 | 0 | 7,3 |
|
Trichoptera |
9,7 | 2,6 | 6,1 |
Была найдена северная граница ареала для вида дафнии D. curvirostris.
Было начато изучение взаимоотношений между видами. Корреляционный анализ наших данных показывает, что некоторые виды чаще встречаются вместе, чем отдельно. Отрицательных корреляций между видами не выявлено (табл. 3).
| Polyphemus pediculus | Ceriodaphnia quadrangula | Scapholeberis mucronata | Daphnia magna | Chydorus sphaericus | Daphnia longispina | Daphnia pulex | |
| Bosmina longispina s.l. | 0,000073 | 0,234147 | 0,000000 | 0,907033 | 0,225548 | 0,000392 | 0,705733 |
| Polyphemus pediculus | 0,721432 | 0,001314 | 0,307055 | 0,160580 | 0,005625 | 0,230308 | |
| Ceriodaphnia quadrangula | 0,127282 | 0,000014 | 0,000605 | 0,307616 | 0,000813 | ||
| Scapholeberis mucronata | 0,850030 | 0,012300 | 0,437491 | 0,842490 | |||
| Daphnia magna | 0,013178 | 0,596885 | 0,025539 | ||||
| Chydorus sphaericus | 0,936694 | 0,019244 | |||||
| Daphnia longispina | 0,893802 |
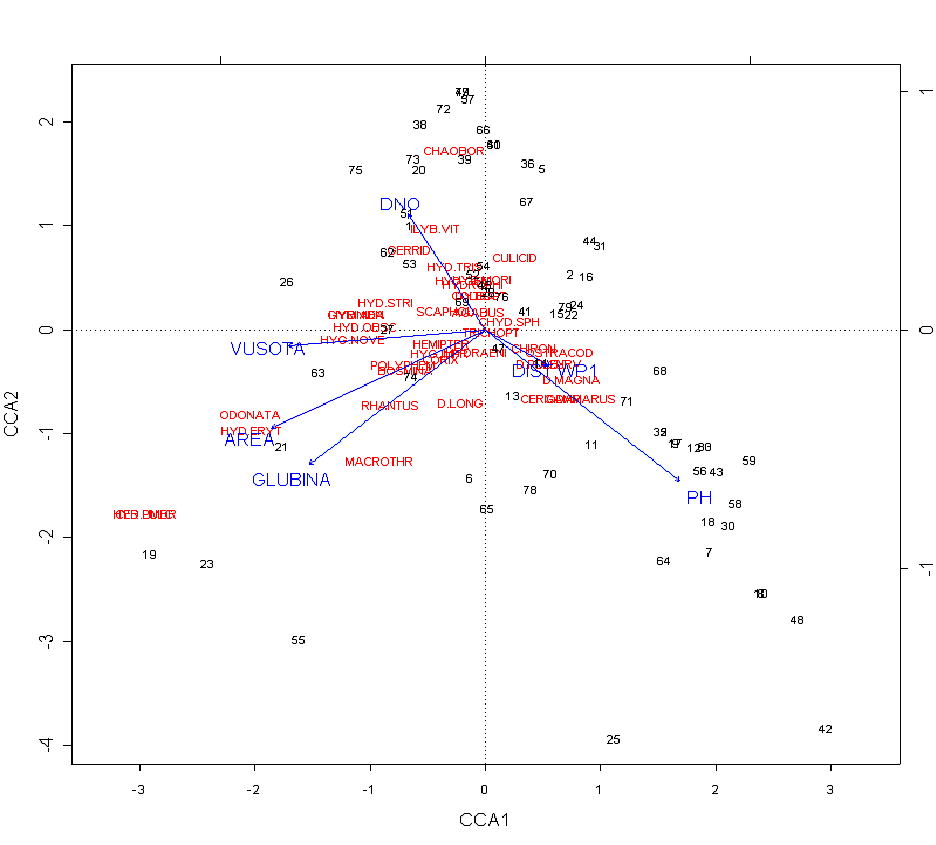
Эти данные совпадают с данными 2002 г, и показывают, что в отдельную группу видов, часто встречающихся совместно, попадают Bosmina, Polyphemus и Daphnia longispina. Эти виды приурочены к наиболее крупным лужам, где чаще встречаются и другие (кроме Polyphemus) хищники. Анализ соответствий подтвердил, что распространение этих видов сильно коррелирует с глубиной и площадью лужи (рис. 1).
числа соответствуют номерам луж, синим цветом обозначены абиотические факторы: AREA - площадь лужи; GLUBINA - глубина лужи; VUSOTA - высота лужи над уровнем моря; DNO - тип дна лужи; DIST WP - электропроводность лужи; PH - уровень рН в луже, красным цветом обозначены названия видов беспозвоночных животных: D_MAGNA -- Daphnia magna; CHYD_SPH -- Chydorus sphaericus; CERIODAP -- Ceriodaphnia quadrangula; SCAPHOL -- Scapholeberis mucronata; D_PULEX -- Daphnia pulex; BOSMINA -- Bosmina longirostris; MACROTHR -- Macrothrix hirsuticornis; CER_PULC -- Ceriodaphnia pulchinella; D_LONG -- Daphnia longispina; POLYPHEM -- Polyphemus pediculus; D_CURV -- Daphnia curvirostris; CULICID – Culicidae; ODONATA – Odonata; OSTRACOD – Ostracoda; COLEOPT – Coleoptera; DYTISC – Ditiscidae; HYG_IMPR -- Hygrotus impressopunctatus; HYG_NOVE -- Hygrotus novemlineatus; HYD_ERYT -- Hydroporus erythrocephalus; HYD_GENI -- Hydroporus geniculatus; HYD_MORI -- Hydroporus morio; HYD_OBSC -- Hydroporus obscurus; HYD_STRI -- Hydroporus striola; HYD_TRIS -- Hydroporus tristis; HYD_UMBR -- Hydroporus umbrosus; AGABUS -- Agabus thomsoni; LIYB_AEN -- IIybius aenescens; ILYB_VIT -- llybius vittiger; RHANTUS -- Rhantus suturellus; GYRINIDA -- Gyrinus opacus; HYDROPHI – Hydrophilidae; HYDRAENI – Hydraenidae; HEMIPTER – Hemiptera; CORIX – Corixidae; GERRID – Gerridae; TRICHOPT -- Trichoptera
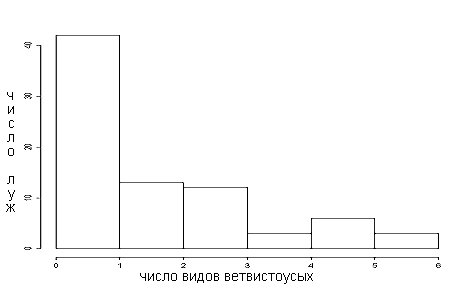
По нашим данным, в луже присутствует от 0 до 6 видов ветвистоусых (рис. 2). Процент луж, не содержащих ни одного вида, возможно, занижен, так как выбирались лужи, где зоопланктон присутствовал.
Позвоночные хищники в обследованных нами водоемах не отмечены (в единственной луже на о.Черепаха в 2003 г. найдены многочисленные мальки колюшки ). Однако в наиболее крупных лужах отмечены крупные личинки неравнокрылых стрекоз, тип хищничества которых (зрительное обнаружение добычи и потребление более крупных кладоцер) сходен с хищничеством позвоночных. Из других беспозвоночных хищников присутствуют Polyphemus pediculus, личинки хаоборусов, клопы-кориксиды, жуки-плавунцы.
Были изучены зависимости числа видов ветвистоусых и насекомых в луже в от некоторых абиотических факторов. Выявлена положительная зависимость (p<0,05) числа видов ветвистоусых в луже от её высоты над уровнем моря, площади и глубины. В ходе работы была выявлена положительная зависимость (p=0) числа видов насекомых в луже от её площади, глубины и высоты над уровнем моря, а также отрицательная зависимость (p=0) числа видов насекомых в луже от уровня pH (чем ниже рН, тем больше число видов жуков).
По результатам канонического анализа соответствий (рис. 1) были выявлены многие зависимости и взаимодействия между различными абиотическими факторами и видами беспозвоночных. Например, наличие в лужах вида Daphnia magna связана с ее электропроводностью и уровнем pH воды, а Daphnia pulex -- только с электропроводностью. Наличие в лужах Daphnia longispina, Hydroporus erythrocephalus, Bosmina longirostris, Coryxidae, Odonata связано с площадью и глубиной лужи. Наличие Odonata и Rhantus suturellus связано с площадью лужи, а Hydroporus striola и Hydroporus obscurus -- с высотой лужи над уровнем моря. Площадь и глубина лужи взаимосвязаны. Некоторые лужи сгруппированы, так как имеют похожие значения некоторых параметров, так, например, лужи №№ 9, 12, 17, 18, 30, 43, 56, 58, 59 имеют значения уровня pH выше среднего (щелочные лужи) , лужи №№ 15, 22, 24, 79 имеют низкую солёность.Наши данные большей частью совпадают с литературными.
Распределение и число видов зависит от электропроводности и pH, но не так сильно, как можно предполагать. В солоноватых лужах могут обитать лишь немногие виды (например, Daphnia magna встречена при солености 4,5 г/л, а Ceriodaphnia dubia и Chydorus sphaericus – при 3,5 г/л). То, что D. magna – наиболее солевыносливый из изученных видов, совпадает с литературными данными.
На основании анализа данных можно сделать вывод, что количество видов ветвистоусых сильнее зависит от глубины лужи, чем от её площади.
Межвидовая конкуренция, судя по нашим данным, не влияет на распределение видов, на что указывает наличие лишь положительных корреляций между встречаемостями пар видов. Видимо, они чаще совместно встречаются там, где для этого есть подходящие уловия. В то же время эти данные нуждаются в уточнении; возможно, результат был бы иным, если анализировались бы не все лужи, а лишь с теми абиотическими условиями, где данный вид может существовать.
Для количественного анализа влияния хищничества на распределение видов данных у нас недостаточно. На качественном уровне можно отметить, что в более крупных и глубоких лужах, где чаще встречаются хищники, чаще встречаются мелкие виды ветвистоусых, чем крупные, а из дафний присутствует D. longispina, что совпадает с литературными данными.
В заключение подчеркнем, что данные, полученные в 2001 и 2002 г.г., носят предварительный характер. В 2003 году проводилась повторная работа.
Главная перспектива данной работы – в накоплении данных многолетних наблюдений. Необходимо исследовать такие факторы, как количество растворенной органики и содержание кислорода. Ценная информация может быть получена при посещении одних и тех же луж с интервалом в 3-4 недели; это позволит сравнить внутрисезонные изменения фауны с межгодовыми.
Другое перспективное направление развития данной работы – изучение морфологической изменчивости, характеристик роста и размножения и генетической структуры популяций ветвистоусых в зависимости от положения лужи и различных факторов среды. Изучение морфологической изменчивости и популяционных характеристик представляется особенно интересным в лужах с разным составом хищников. Изучение генетической изменчивости позволило бы яснее представить характеристики распространения видов и степень обмена между лужами одного острова и разных островов.
Мы благодарим Е. Альтшулера, П. Волкову и А.Б. Шипунова за помощь в статистической обработке материалов, А.Н. Квашенко и В.И. Люляева за помощь в доставке группы на острова, администрацию Кандалакшского заповедника за разрешение на работу на территории заповедника.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |