Поведение ловчих листьев насекомоядных растений из рода Drosera в лабораторных условиях активно изучалось еще в конце 19 века (Kellerman, Raumer, 1878; Busgen, 1883; Darwin, 1903). Эти исследования продолжаются до сих пор с применением современных методов (Chandler, Anderson, 1976; Hanslin, Karlsson, 1996; Муравник, 2002). К сожалению, в лабораторных условиях невозможно учесть влияние погодных факторов на поведение листьев, а также оценить число доступных для растения в естественных условиях беспозвоночных. Кроме того, на успех ловли беспозвоночных в естественных условиях могут влиять и другие факторы, например, тип добычи, клептопаразитизм муравьев (Thum, 1989a) и условия обитания (Thum, 1989a; Gomes, 1998).
Нами были впервые проведены непрерывные наблюдения за поведением ловчих листьев Drosera rotundifolia в естественных условиях в Средней России и в районе Полярного круга (Volkova, Shipunov, 2005, см. также табл. 1). Результаты этих наблюдений свидетельствуют о том, что изменение состояния листовой пластинки у D. rotundifolia, по-видимому, носит случайный характер и лишь корректируется внешними факторами, такими как относительная влажность воздуха, атмосферное давление и наличие насекомых на листовой пластинке. Наблюдаемое увлажнение листовой пластинки, ее заворачивание, а также загибание краевых волосков через некоторое время после попадания насекомого на листовую пластинку можно рассматривать как совпадение (Volkova, Shipunov, 2005), что противоречит данным о поведении ловчих листьев D. rotundifolia в лабораторных условиях (Kellerman, Raumer, 1878; Busgen, 1883; Darwin, 1903). Такое несоответствие полученных в полевых условиях результатов лабораторным данным подтверждает необходимость дальнейшего изучения поведения ловчих листьев насекомоядных растений в естественных условиях.
|
Таблица 1. Поведение ловчих листьев Drosera rotundifolia (Volkova, Shipunov, 2005) и D. anglica в Северной Карелии: доля листьев с заметными изменениями (общее число листьев, за которыми велось наблюдение, было принято за 100%). |
|||||||
|
Характеристика листовой пластинки |
Изменения после попадания добычи на лист в течение... |
Изменения в отсутствие добычи на листе |
Изменения при постоянном числе беспозвоночных на листе |
Не отмеченное ранее в литературе поведение |
|||
|
< 1 h |
1-2 h |
3 h |
|||||
|
степень увлажненности |
D. rotundifolia |
0 |
0 |
17 |
67 |
63 |
88 |
|
D. anglica |
22 |
11 |
0 |
100 |
43 |
40 |
|
|
форма |
D. rotundifolia |
0 |
17 |
17 |
83 |
100 |
25 |
|
D. anglica |
56 |
0 |
0 |
50 |
29 |
80 |
|
| доля загнутых краевых железистых волосков |
D. rotundifolia |
0 |
50 |
50 |
100 |
75 |
50 |
|
D. anglica |
13 |
13 |
13 |
91 |
14 |
78 |
|
В экспериментах с искусственным помещением насекомых на листья D. rotundifolia и D. intermedia в полевых условиях, были обнаружены существенные межвидовые различия в доле насекомых, остающихся на листьях через 24 часа (Thum, 1988; Thum, 1989a; Thum, 1989b). Болотные муравьи утаскивали две трети насекомых с листьев D. rotundifolia, тогда как насекомые на листьях D. intermedia оставались практически нетронутыми (Thum, 1989b). Такие различия могут быть объяснены разницей в морфологии растений (листья D. rotundifolia распростерты по субстрату, а листья D. intermedia подняты почти вертикально) или приуроченностью этих видов к различным микробиотопам (Thum, 1988; Thum, 1989a). Предполагается, что D. rotundifolia ловит в три раза больше насекомых по сравнению с D. intermedia, так как эти два вида имеют одинаковые значения биомассы переваренных насекомых, отнесенной к биомассе растения (Thum, 1989a; Thum, 1989b). Надо отметить, что в работах M. Thum (1988, 1989a, 1989b) наблюдения за состоянием растений проводились не чаще одного раза в сутки, что не позволяет подробно описывать поведение ловчих листьев.
Все вышеизложенное позволяет предположить существование различных стратегий ловли насекомых у представителей рода Drosera, различающихся по морфологии и микробиотопической приуроченности.
На севере Европейской России наряду с D. rotundifolia произрастает D. anglica -- вид, сходный с D. intermedia по морфологии и микробиотопической приуроченности (Webb, 1993). Мы решили провести наблюдения за поведением ловчих листьев D. anglica в естественных условиях, чтобы сопоставить его с поведением ловчих листьев D. rotundifolia и выявить возможные стратегии ловли насекомых.
Наблюдения велись за двумя растениями D. anglica в Лоухском районе республики Карелия, находящимися в естественных условиях (сфагновая сплавина на северном берегу оз. Ближнее в 200 м к северу от дер. Нижняя Пулонга; 66 град. 18,5' с.ш., 33 град. 7,5' в.д.) с 16 по 18 июля 2005 года. Растения были выбраны случайным образом из типичной для данных природных условий популяции. Непрерывные наблюдения за парой исследуемых растений продолжались в течение 52 часов. На каждом из растений было по 7 ловчих листьев, причем за время наблюдений один лист засох, и один лист развернулся. В течение всего срока наблюдений для каждого ловчего листа один раз в 40 минут визуально оценивали по разработанной ранее методике форму листовой пластинки, степень ее увлажненности липкой слизью и долю загнутых краевых железистых волосков (Volkova, Shipunov, 2005). Мы также подсчитывали число насекомых, находящихся на листовой пластинке, и визуально оценивали их состояние (свежие, полупереваренные, хитиновый скелет). Один раз в 20 минут регистрировали температуру и относительную влажность воздуха при помощи электронного термометра и электронного психрометра. Один раз в два часа измеряли атмосферное давление при помощи бытового барометра.
Более половины листьев реагировали на пойманное насекомое менее чем через час после попадания насекомого на листовую пластинку (табл. 1). Почти у 60% листьев за этот период времени листовая пластинка заворачивалась, у нескольких листьев наблюдалось увлажнение листовой пластинки или увеличение доли загнутых краевых железистых волосков. Реакция листьев на пойманное насекомое в течение двух-трех часов с момента попадания насекомого на листовую пластинку практически отсутствовала. При этом у всех листьев в отсутствии насекомых на листовой пластинке наблюдались не имеющие четкой периодичности изменения степени ее увлажненности, или доли загнутых краевых волосков, или, реже, формы продолжительностью 1-6 часов. Такое поведение было отмечено для половины исследованных листьев и в присутствии на их листовой пластинке постоянного числа непереваренных насекомых. Часто наблюдалось отличное от описанного в литературе поведение листа при наличии на его пластинке недавно пойманного насекомого. Так, 40% листьев не выделяли слизи при попадании на них насекомых, листовая пластинка 80% листьев не заворачивалась, а краевые волоски 80% листьев не были загнутыми при нахождении на листовой пластинке непереваренного насекомого (табл. 1).
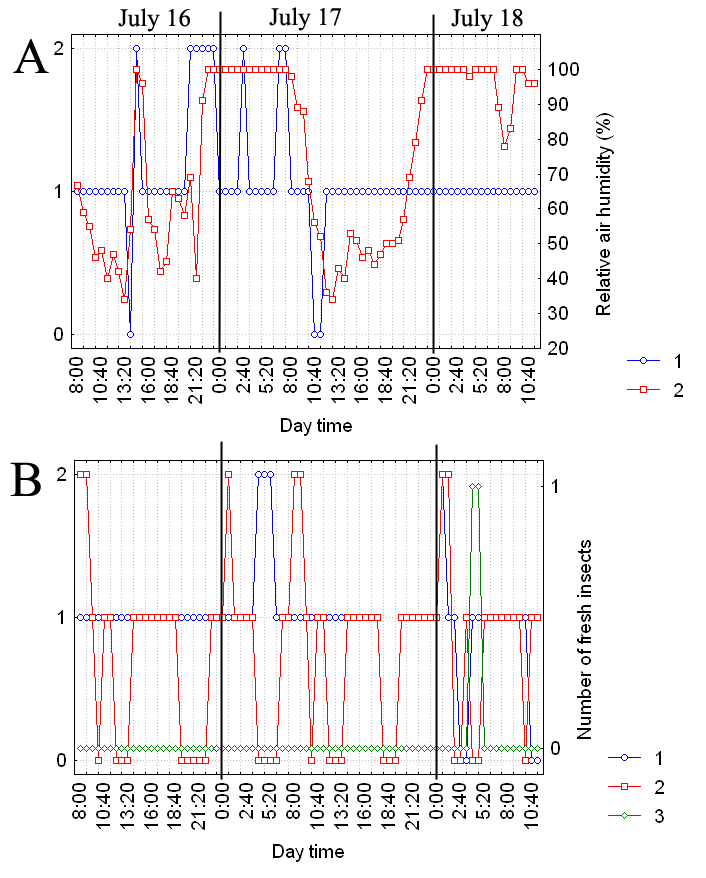
Как видно из вышеизложенного, изменения различных характеристик листовой пластинки происходят несогласовано. При этом наблюдается четкая связь степени увлажненности листовой пластинки с относительной влажностью воздуха (рис. 1).
A: 1 -- степень увлажненности листовой
пластинки, 2 -- относительная влажность
воздуха (%);
B: 1 -- форма листовой пластинки, 2 -- доля
загнутых волосков, 3 -- число свежих
насекомых на листовой пластинке.
В период наблюдений температура воздуха в среднем оставалась постоянной и составляла 22 град. C. Суточные колебания температуры воздуха были хорошо выражены: температура достигала своего минимального значения в 3-4 утра, опускаясь до 10-11 град. С, и своего максимального значения в 12-13 дня, поднимаясь до 32-38 град. С. Атмосферное давление в первые сутки наблюдений составляло 757-758 мм рт. ст., а потом стало повышаться, достигнув 760 мм рт. ст. Заметного влияния температуры воздуха и атмосферного давления на характеристики листовой пластинки не было обнаружено.
Число пойманных насекомых варьировало от 0 до 1,4 насекомых на лист в сутки, составляя в среднем 0,6 насекомых на лист в сутки. Мы не обнаружили зависимости этого параметра от погодных условий.
Наши наблюдения показывают, что различные характеристики листовой пластинки D. anglica в Карелии изменяются практически независимо друг от друга и от наличия насекомых на листе, а степень увлажненности листовой пластинки в значительно мере зависит от относительной влажности воздуха. Эти наблюдения согласуются с результатами для D. rotundifolia в этом регионе (Volkova, Shipunov, 2005). Подробное обсуждение этих фактов можно найти в нашей статье, посвященной поведению D. rotundifolia (Volkova, Shipunov, 2005). Здесь же мы хотели бы обратить внимание на существенные различия в поведении ловчих листьев D. anglica и D. rotundifolia, проявляющиеся несмотря на сходство поведения ловчих листьев этих двух видов росянки в общих чертах.
У D. anglica отмечена быстрая реакция (в течение 1 часа) на попадание насекомого на листовую пластинку, в отличие от медленной реакции (в течение 2-3 часов) у D. rotundifolia. Интересно, что при попадании насекомого на листовую пластинку у D. anglica чаще всего происходит заворачивание листовой пластинки, тогда как у D. rotundifolia удержание добычи происходит преимущественно за счет краевых волосков(табл. 1). Изменение положения краевых волосков в отсутствии насекомых на листовой пластинке характерно практически для всех ловчих листьев обоих видов росянки. При этом изменение степени увлажненности ловчих листьев D. anglica происходит чаще, а формы листовой пластинки -- реже по сравнению с D. rotundifolia (табл. 1). Изменение состояния листовой пластинки при наличии на ней постоянного числа насекомых происходит у D. anglica значительно реже, чем у D. rotundifolia (в особенности это касается формы листовой пластинки и положения краевых волосков). Отличное от описанного в литературе поведение при попадании насекомого на листовую пластинку наблюдалось у D. anglica в целом чаще, чем у D. rotundifolia (хотя для степени увлажненности листовой пластинки наблюдается обратная закономерность).
Интенсивность ловли насекомых у D. rotundifolia в Карелии составляет в среднем 0,3 насекомых на лист в сутки, варьируя от 0 до 0,7 насекомых на лист в сутки (Волкова, Шипунов, неопубл. данные). Таким образом, наши данные не подтверждают предположения M. Thum (1989a) о большем числе пойманных насекомых у D. rotundifolia. Скорее всего, одинаковые значения биомассы пойманных насекомых, отнесенной к биомассе растения (Thum, 1989b) объяснялись не межвидовыми различиями в числе пойманных насекомых (Thum, 1989a), а существенно меньшей биомассой растений D. rotundifolia по сравнению с D. intermedia, а также межвидовыми различиями в биомассе пойманных насекомых.
Отмеченные различия в поведении ловчих листьев у D. anglica и у D. rotundifolia свидетельствуют о разных стратегиях ловли насекомых. Такая разница может быть объяснена различиями в морфологии и микробиотопических предпочтениях (Webb, 1993; Волкова, 2002), а следовательно и в типе добычи этих двух видов росянки (Thum, 1989a; Thum, 1989b). Слабая зависимость поведения ловчих листьев от наличия на них насекомых позволяет рассматривать ловушки росянки как переходные структуры между активными и пассивными листьями-липучками, вопреки распространенному пониманию листьев росянки как активных листьев-липучек (Williams, 1976).
Сбор данных для настоящей работы проводился в ходе Беломорской экспедиции Московской Гимназии на Юго-Западе, проходившей под руководством С. Глаголева. В полевых наблюдениях неоценимую помощь оказали Ю. Быков, М. Левина и К. Марквичева.
Волкова П.А. 2002. О морфологических отличиях трех видов росянки (Drosera L., Droseraceae) // Международная научная конференция по систематике высших растений: Тез. докл. -- М.: Изд-во Центра охраны дикой природы. 29-30.
Муравник Л.Е. 2002. Влияние химической стимуляции на ультраструктуру секреторных клеток железистых волосков двух видов Drosera. Физиол. раст. 47 (4): 614-623.
Busgen M. 1883. Die bedeutung des Insektenfangs fur D. rotundifolia. Bot. Zeitung 41: 569-577.
Chandler G.E., Anderson J.W. 1976. Studies on the nutrition and growth of Drosera species with reference to the carnivorous habit // New Phytol. 76: 129-141.
Gomez L.D. 1998. Natural history and occurence of the "insectivorous plant" Drosera capillaris (Droseraceae) in Costa Rica // Revista de Biologia Tropical. 46 (4): 1033-1037.
Kellerman C., Rauner E. 1878. Vegetations versache an D. rotundifolia mit und ohne Fleishfutterung. Bot. Zeitung 36: 209-218.
Hanslin H.M., Karlsson P.S. 1996. Nitrogen uptake from prey and substrate as affected by prey capture level and plant reproductive status in four carnivorous plant species. Oecologia (Berlin) 106: 370-375.
Thum M. 1988. The significance of carnivory for the fitness of Drosera in its natural habitat 1. The reactions of D. intermedia and D. rotundifolia to supplementary feeding. Oecologia 75: 472-480.
Thum M. 1989a. The significance of opportunistic predators for the simpatric carnivorous plant species Drosera intermedia and Drosera rotundifolia. Oecologia 81: 397-400.
Thum M. 1989b. The significance of carnivory for the fitness of Drosera in its natural habitat 2. The amount of captured prey and its effect on Drosera intermedia and Drosera rotundifolia. Oecologia 81: 401-411.
Webb D.A. 1993. Drosera L. // Flora Europaea. Vol. 1. Ed. 2. University Press, Cambridge: 421-422.
Williams S.E. 1976. Comparative sensory physiology of the Droseraceae – the evolution of plant sensory system. Proceedings of the American Philosophical society. 120: 187-204.
Volkova P.A., Shipunov A.B. 2005. The behaviour of Drosera rotundifolia L. (Droseraceae) trapping leaves in natural habitats // Carnivorous plant newsletter 34 (1): 7-15.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |