У некоторых в норме обоеполых цветков Geranium sylvaticum L. пыльники всех или нескольких из десяти имеющихся тычинок редуцируются, и такие цветки становятся женскими или "промежуточными" соответственно. Это препятствует самоопылению растений герани, позволяет женским цветкам продуцировать больше семян или семена с большим запасом питательных веществ, что благотворно сказывается на развитии популяции в целом (Asikainen, Mutikainen, 2003; Ramula, Mutikainen, 2003). Отмечена отрицательная связь доли женских цветков в популяции с семенной продуктивностью обоеполых цветков (Asikainen, Mutikainen, 2003). Отсюда следует, что соотношение полов в популяции заслуживает детального изучения, так как является важной характеристикой репродуктивного статуса составляющих ее растений.
Частота встречаемости женских цветков была подробно изучена на 23 популяциях герани в Финляндии (Asikainen, Mutikainen, 2003). Доля женских цветков в популяции по данным этих авторов варьировала от 0,4 до 27,2% (составляя в среднем 11,5%) и зависела от условий обитания. К примеру, женские цветки реже встречаются при более благоприятных условиях обитания, в частности в южной Финляндии по сравнению с северной частью страны (Asikainen, Mutikainen, 2003). “Промежуточные” цветки были обнаружены менее чем в половине исследованных популяций в очень небольшом количестве (менее 1%). Лишь в одной из популяций, расположенных на юге страны, доля “промежуточных” цветков составляла 42% (Asikainen, Mutikainen, 2003).
G. sylvaticum распространена практически повсеместно на территории Евразии. Поэтому мы решили проверить данные E. Asikainen и P. Mutikainen (2003) о влиянии географической широты местности на соотношение полов в популяции герани лесной, полученные на финских популяциях, на более обширной территории Европейской России.
Работу проводили в июне-июле 2003-2005 годов в Удомельском районе Тверской области -- 58° 15' с.ш., 34° 30' в.д. (564 растения из 21 популяции) и в Североморском районе Мурманской области -- 69° 10' с.ш., 36° 00' в.д. (274 растения из четырех популяций). Популяцией мы называли топографически обособленную группу растений. Мы исследовали по одному цветку у каждого цветущего растения из популяции. Измерялись такие морфологические параметры растений, как максимальный диаметр наибольшего прикорневого листа, расстояние от места крепления черешка до угла ближайшего выреза на листе, длина лепестка, ширина лепестка и длина чашелистика, кроме того мы отмечали число фертильных тычинок и цвет венчика цветка (по четырехбалльной визуальной шкале). Чрезвычайно важно было исследовать только что распустившиеся цветки, так как на более поздних стадиях развития цветка пыльники легко отделяются от тычиночных нитей (личные наблюдения), что может привести к ошибке при подсчете числа фертильных тычинок.
Впоследствии по модифицированным шкалам Элленберга (Hill et al., 1999) мы вычислили степень освещенности и степень влажности местообитания, степень кислотности почвы и содержание азота в почве на основании видового списка массово произрастающих в пределах популяции высших растений.
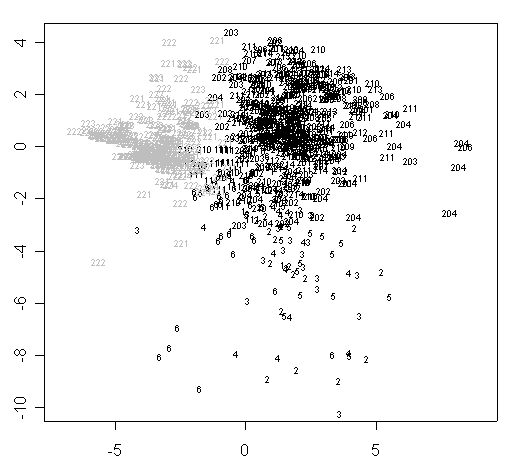
В ходе обработки морфологических данных методом многомерного шкалирования было выявлено существование двух географических морфотипов герани лесной (рис. 1). Различия этих морфотипов по отдельным морфологическим характеристикам подтверждаются результатами теста Стьюдента (p<<0,05).
Рисунок 1. Классификация исследованных растений на основании их морфологии методом многомерного шкалирования (Kruskal’s non-metric MDS, daisy metric)
Условные обозначения: Число обозначает растение из популяции с соответствующим номером (согласно табл. 2). Черным цветом обозначены популяции из Тверской области, серым цветом -- из Мурманской области.
Для Мурманской области характерны растения с относительно мелкими листьями и крупным венчиком (табл. 1). Вероятно, существование двух морфотипов вызвано значительными различиями в условиях обитания. Бесспорный факт существования таких различий подтверждается и разницей в значениях некоторых эдафических факторов, рассчитанных по шкалам Элленберга. В Мурманской области отмечено относительно низкое содержание азота в почве (2-3 по шкале Элленберга в Мурманской области: низкая минерализация и 3,5-7,5 в Тверской области: умеренная и высокая минерализация) и относительно высокая кислотность почвы (3-5: умеренно кислые почвы и 5-7, 5: слабо кислые и нейтральные почвы соответственно).
Таблица 1. Квартильный размах морфологических характеристик
|
характеристика |
Тверская область |
Мурманская область |
|
максимальный диаметр листа (см) |
123-163 |
40-52 |
|
относительная глубина разрезанности листа (%) |
83-89 |
70-76 |
|
длина лепестка (мм) |
8-13 |
14-17 |
|
ширина лепестка (мм) |
5-8 |
8-10 |
|
длина чашелистика (мм) |
10-12 |
8-9 |
Мы не обнаружили ни одного женского цветка в исследованных популяциях Тверской обл. В Мурманской обл. в одной из четырех популяций был найден единственный женский цветок (0,9% от общего числа цветков в популяции), в прочих трех популяциях женских цветков не было обнаружено. Наши результаты свидетельствуют о большей редкости женских цветков в популяции, чем это указывалось E. Asikainen и P. Mutikainen (2003).
Доля обоеполых цветков в популяции в Тверской области в среднем ниже (17-100 (57+/-6) %), чем на Баренцевом море при более жестких климатических условиях (96-100 (97+/-0,5) %) в противоположность данным E. Asikainen и P. Mutikainen (2003) (табл. 2). Большая вариация доли обоеполых цветков в Тверской обл. не объясняется топологией популяций или эдафическими факторами.
Таблица 2. Доля растений с различным числом фертильных тычинок по популяциям
|
регион |
номер популяции |
число растений |
число фертильных тычинок |
|||||||||
|
0 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|||
|
Тверская область |
1 |
6 |
0 |
0 |
0 |
0 |
0 |
16.7 |
16.7 |
16.7 |
16.7 |
33.3 |
|
2 |
18 |
0 |
5.6 |
0 |
5.6 |
5.6 |
16.7 |
5.6 |
16.7 |
27.8 |
16.7 |
|
|
3 |
20 |
0 |
0 |
0 |
0 |
0 |
20 |
15 |
15 |
25 |
25 |
|
|
4 |
16 |
0 |
0 |
0 |
0 |
0 |
18.8 |
12.5 |
12.5 |
25 |
31.2 |
|
|
5 |
18 |
0 |
0 |
0 |
0 |
0 |
16.7 |
11.1 |
16.7 |
22.2 |
33.3 |
|
|
6 |
40 |
0 |
0 |
2.5 |
5 |
5 |
2.5 |
0 |
7.5 |
15 |
62.5 |
|
|
111 |
17 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
11.8 |
29.4 |
58.8 |
|
|
201 |
26 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
7.7 |
11.5 |
80.8 |
|
|
202 |
36 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
5.6 |
8.3 |
86.1 |
|
|
203 |
28 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
14.3 |
0 |
85.7 |
|
|
204 |
59 |
0 |
0 |
0 |
0 |
0 |
0 |
1.7 |
5.1 |
23.7 |
69.5 |
|
|
205 |
14 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
21.4 |
78.6 |
|
|
206 |
31 |
0 |
0 |
0 |
0 |
0 |
0 |
3.2 |
3.2 |
16.1 |
77.4 |
|
|
207 |
18 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
33.3 |
66.7 |
|
|
208 |
9 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
|
209 |
8 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
12.5 |
87.5 |
|
|
210 |
61 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
3.3 |
8.2 |
88.5 |
|
|
211 |
33 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
|
212 |
28 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
|
213 |
23 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
8.7 |
91.3 |
|
|
214 |
61 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
|
Мурманская область |
221 |
108 |
0.9 |
0 |
0 |
0 |
0 |
0 |
0.9 |
0 |
1.9 |
96.3 |
|
222 |
108 |
0 |
0 |
0.9 |
0.9 |
0 |
0 |
0 |
0 |
0 |
98.1 |
|
|
223 |
41 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
|
224 |
18 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
100 |
|
Доля "промежуточных" цветков в популяции колеблется от 9 до 84% в Тверской области (43+/-6% в среднем) и от 0 до 4% (1,5+/-1%) в Мурманской обл. (табл. 2). Таким образом, данные E. Asikainen и P. Mutikainen (2003) о низкой доли "промежуточных" цветков в популяции согласуются с нашими данными только для Мурманской обл.
Таким образом, наши данные о доле женских цветков в различных популяциях герани в Европейской России существенно не согласуются с данными E. Asikainen и P. Mutikainen (2003), полученными в Финляндии. Возможно, такие расхождения связаны с различием в размерах исследованных популяций (размер исследованных популяций в Европейской России был на один-два порядка меньше размера исследованных популяций в Финляндии). Можно предположить и существование зависимости доли женских цветков в популяции от географической широты местности. Наконец, расхождения в половом составе популяций можно объяснить методической ошибкой при подсчете числа фертильных тычинок. В работах P. Mutikainen с соавторами (Asikainen, Mutikainen, 2003; Ramula, Mutikainen, 2003) не указана фаза цветения, на которой проводилось определение пола цветка. Если определение пола проводилось на поздних стадиях цветения, то пыльники фертильных тычинок могли самопроизвольно отделяться от тычиночных нитей. В результате такой ошибки обоеполые цветки могли быть приняты за “промежуточные”, а “промежуточные” – за женские.
Asikainen E., Mutikainen P. Female frequency and relative fitness of females and hermaphrodites in gynodioecious Geranium sylvaticum (Geraniaceae) // American Journal of Botany, 2003, 90: 226-234.
Hill M.O., Mountford J.O., Roy D.B., Bunce R.G.H. Ellenberg's indicator values for British plants // ECOFACT, Technical Annex. Vol. 2. 1999.
Ramula S., Mutikainen P. Sex allocation of females and hermaphrodites in the gynodioecious Geranium sylvaticum // Annals of Botany, 2003, 92: 207-213.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |