
Форма и размер листовых пластинок и пыльцевых зерен являются важными диагностическими признаками во многих группах растений. Проведены количественные исследования изменений размеров и формы листовых пластинок при гербаризации, а также размеров пыльцевых зерен после ацетолизной обработки и в условиях высокого вакуума при СЭМ у Nymphaea alba, N. candida и N. tetragona в Европейской России. При гербаризации длина листовой пластинки уменьшается в 1,05-1,4 раза, а ширина -- в 1,15-1,5 раза. Степень деформации листовой пластинки в продольном направлении не связана достоверно со степенью ее деформации в поперечном направлении. Достоверные различия между размерами пыльцевых зерен одного и того же цветка до и после ацетолиза отсутствуют. Условия высокого вакуума, необходимого для СЭМ, приводят к заметной деформации пыльцевых зерен (значения малого экваториального диаметра существенно уменьшаются, тогда как значения большого экваториального диаметра остаются неизменными).
В эпоху бурного развития молекулярной систематики использование морфологических признаков для диагностики видов остается по-прежнему актуальным. Хорошо известно, что систематики растений работают в основном не с живым материалом, а с гербарными коллекциями. Как правило, в результате исследований гербарных образцов вырабатываются рекомендации по диагностике отдельных видов и на живом материале. Преимущества гербария перед живыми растениями для работы систематика очевидны и неоспоримы. Работа в гербарии позволяет существенно расширить географический охват исследований, сводя при этом к минимуму материальные и временные затраты. Кроме того, изучение гербарного материала позволяет провести консультацию со специалистами по группе по истечении полевого сезона и даже через несколько десятков и сотен лет после сбора живого материала. Однако хорошо известно, что многие основные диагностические признаки изменяются при гербаризации, что может сделать невозможным широко практикующийся в настоящее время прямой перенос результатов гербарных исследований на живой материал. Несмотря на это, количественные исследования изменений растительного материала при гербаризации практически отсутствуют в литературе (см. Vermeulen, 1947).
Морфология и размеры пыльцевых зерен также часто используются для диагностики видов растений. Исследования палиноморфологии нередко проводят после ацетолизной обработки пыльцы или при помощи сканирующей электронной микроскопии (СЭМ), проводимой в условиях глубокого вакуума. Эти процедуры могут изменять размеры пыльцевых зерен, хотя результаты таких исследований часто используются впоследствии для сравнения с измерениями интактных пыльцевых зерен. В литературе практически отсутствуют данные количественных исследований формы и размера пыльцевых зерен после ацетолиза или СЭМ.
В процессе подробного изучения морфологической изменчивости кувшинок (Nymphaea candida, N. alba и N. tetragona) в Европейской России (Volkova, Shipunov, в печати) мы пришли к необходимости количественного изучения изменений диагностических признаков, используемых в этой таксономической группе, при традиционных способах обработки живого материала.
Род Nymphaea широко распространен в пресных водоемах и водотоках зоны тропического и умеренного климата обоих полушарий. Принято считать, что в России произрастает три вида кувшинки: N. alba L., N. candida J. Presl и N. tetragona Georgi, которые различаются в основном деталями строения цветка, а также морфологией листьев и пыльцевых зерен (Крупкина, 2001). N. alba распространена в Европейской части России, N. candida – в Европейской части России и в Сибири, N. tetragona – в Сибири, на Дальнем Востоке и на Кольском полуострове (Комаров, 1937; Muntendam et al., 1996). Все эти виды характеризуются значительной морфологической изменчивостью, что позволяет разным систематикам выделять на территории России от одного до 11 видов кувшинки (Heslop-Harrison, 1955). Это обстоятельство затрудняет выбор диагностических признаков для разграничения трех традиционно выделяемых российских видов рода Nymphaea.
Многие основные диагностические макроморфологические признаки (например, форма и размер листовой пластинки: Маевский, 1976; Крупкина, 2001; Uotila, 2001) в роде Nymphaea существенно изменяются при гербаризации (Лисицына, 2003). Важными диагностическими признаками в роде Nymphaea считаются также размер и форма пыльцевых зерен (Куприянова, 1976; Muntendam et al., 1996; Uotila, 2001), при этом количественные исследования этих параметров проведены только в двух работах. Л.А. Куприянова (1976) изучала ацетолизированные пыльцевые зерна кувшинок европейской части СССР, а J.B. Muntendam с соавторами (1996) измеряли пыльцевые зерна голландских N. alba s.l. (incl. N. candida) на СЭМ-фотографиях.
Целью настоящей работы являлись количественные исследования изменений размеров и формы листовых пластинок при гербаризации, а также размеров пыльцевых зерен после ацетолизной обработки и в условиях высокого вакуума при СЭМ у представителей рода Nymphaea в Европейской России.
Исследована 41 популяция Nymphaea candida из Республики Карелия, Московской, Тверской, Челябинской, Липецкой, Иркутской и Мурманской областей, 5 популяций N. alba в дельте Волге (Астраханская обл.) и 3 популяции N. tetragona на юго-западном берегу оз. Байкал (Республика Бурятия). Схема расположения исследованных популяций и подробная информация о них даны в нашей статье (Volkova, Shipunov, в печати).
Для исследования обводили на бумаге контур наибольшей листовой пластинки одного растения из каждой популяции, измеряли ее максимальную длину и ширину и затем гербаризировали. Контуры гербаризированных листовых пластинок также обводили на бумаге и измеряли, когда листья были полностью высушены. Кроме того, гербаризировали цветки одного или нескольких растений из каждой популяции для последующего анализа размера пыльцевых зерен.
Для исследования формы листовых пластинок применили геометрическую морфометрию (метод тонких пластин, TPS). Этот подход позволяет исследовать разнообразие форм объектов, исключая размерный фактор (Bookstein, 1991; Adams et al., 2002). Были использованы контуры листовой пластинки N. candida и N. tetragona, заимствованные из «Flora Nordica» (Uotila, 2001) и из «Illustrated Flora of Northern US and Canada» (Britton, Brown, 1913) в качестве «якорных образцов».
Для характеристики формы листовой пластинки методами TPS на ее контуре ставили 100 меток с равными промежутками между ними при помощи экранного дигитайзера tpsDig (Rohlf, 2001b). Начальная метка располагалась в вершине выемки между лопастями листовой пластинки. Координаты эталонной (усредненной) конфигурации, а также значения главных, относительных и частных трансформаций, характеризующие меру отличия образца от эталона, вычисляли при помощи программы tpsRelw, реализующей идею геометрической морфометрии в форме метода, аналогичного методу главных компонент (Rohlf, 2001a). Исходные координаты экземпляров нормировали относительно эталонной конфигурации при помощи прокрустова наложения, коэффициент шкалирования альфа принимали равным 0.
Форму листьев до и после гербаризации сравнивали при помощи программы tpsRegr (Rohlf, 2005). Усреднение формы листовых пластинок проводили при помощи программы tpsSuper (Rohlf, 2003). Редактирование и конвертирование файлов данных производили при помощи вспомогательной программы tpsUtil (Rohlf, 2000).
Для исследования изменений размеров пыльцевых зерен после ацетолизной обработки, провели ацетолиз пыльцы из десяти гербаризированных цветков по стандартной методике (Основы…, 2000). Затем размеры пыльцевых зерен (большой и малый экваториальный диаметр) были сопоставлены с аналогичными показателями для неацетолизированной пыльцы из тех же цветков (по 15-20 пыльцевых зерен из каждого цветка до и после ацетолиза).
Для исследования изменений размеров пыльцевых зерен в условиях высокого вакуума при СЭМ пыльцу прикрепляли к столику для СЭМ при помощи маникюрного лака, напыление проводили в ионнораспылительной установке IB-3 фирмы Eiko сплавом золота и палладия (примерная толщина напыления 25 нм). Микрографии получали при помощи сканирующего электронного микроскопа Camscan S-2 при ускоряющем напряжении 20 кВ в режиме SEI. Оцифровка изображения производилась при помощи компьютерной программы Scan Microcapture 2.20119 (разработчик А.В. Григорьев). Всего была сделана 421 микрофотография. Затем размеры пыльцевых зерен (большой и малый экваториальный диаметр) были сопоставлены с аналогичными промерами пыльцы из тех же цветков, сделанными под световым микроскопом.
Измерения неацетолизированной пыльцы проводили под световым микроскопом МИКМЕД-1 на временных препаратах (первые 15-20 пыльцевых зерен, обнаруженные в подходящем положении – вид точно с полюса). Все измерения проводили при помощи окуляр-микрометра (×15) с точностью до 1 мкм на увеличении 1,5×15×40. Всего было измерено 564 пыльцевых зерна
Для выявления различий и взаимосвязей между выборками метрических данных использовались соответственно параметрический тест Стьюдента и параметрический коэффициент корреляции Пирсона. Тест на соответствие распределения значений данных нормальному был проведен при помощи вспомогательной программы, написанной А.Б. Шипуновым на основе теста Шапиро-Уилкса. Для выявлений различий в форме листовых пластинок использовался генерализованный тест Гудала (generalized Goodall F-test), реализованный в программе tpsRegr (Rohlf, 2005). Для классификации листьев по их форме методом геометрической морфометрии использовали анализ главных компонент, так как в этом случае мы имели дело с непрерывными переменными матрицы относительных деформаций. Для всех вычислений и графического представления результатов использовалась компьютерная статистическая среда R (R Development Core Team, 2004).
При классификации листовых пластинок методом главных компонент на основании их формы (данные геометрической морфометрии) можно выделить три области (рис. 1). Одна из этих областей («зона A») представлена исключительно листьями N. alba до гербаризации. Вторая область («зона B») представлена листьями N. tetragona и гербаризированными листьями растений из других регионов России. В центральной части этой области располагается «якорный образец» N. tetragona. Между «зоной A» и «зоной B» помимо «якорного образца» N. candida находятся все остальные исследованные листовые пластинки, включая несколько гербаризированных.
Рисунок 1. Распределение исследованных растений в пространстве двух первых главных компонент анализа главных компонент относительных деформаций контуров листовой пластинки
Каждый лист обозначен как xx-yy, где xx – порядковый номер популяции, а yy – порядковый номер растения в этой популяции. Индекс h или d указывает на листовые пластинки после гербаризации. Черными точками обозначены «якорные образцы». Отграничены области растений из Астраханской обл. (зона A) и из Республики Бурятия (зона B). Стрелки соединяют одни и те же листовые пластинки до и после гербаризации.
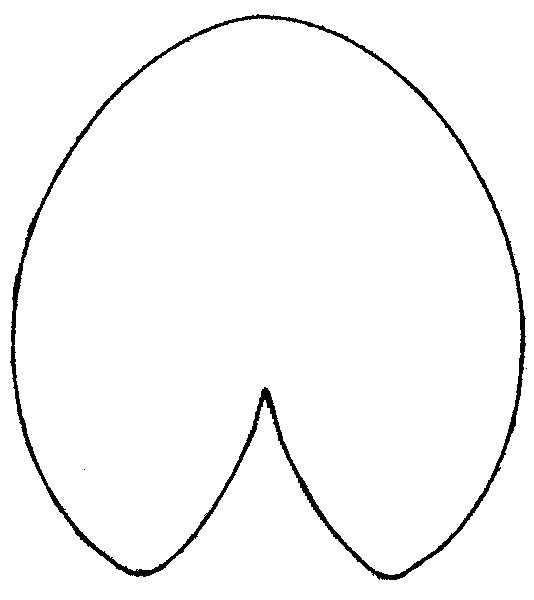
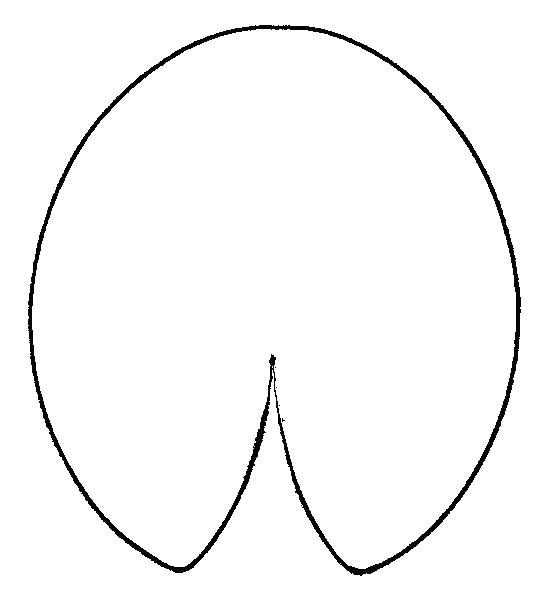
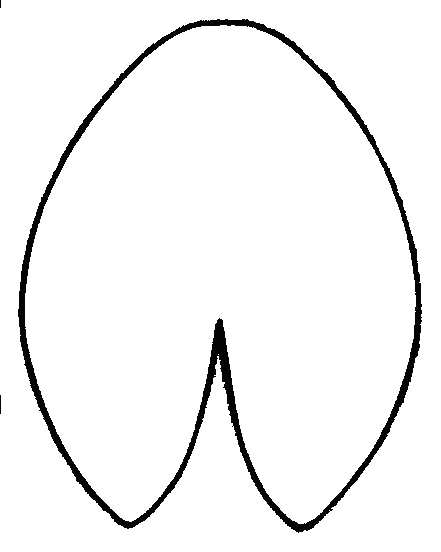
Легко заметить, что после гербаризации листовые пластинки из «зоны A» «перемещаются» в центральную область или даже в «зону B», а листовые пластинки из центральной области «перемещаются» в «зону B» (рис. 1). Различия по форме между одними и теми же листовыми пластинками до и после гербаризации высоко достоверны (generalized Goodall F-test: p=0). При «продвижении» из «зоны A» в «зону B» листовые пластинки становятся более продолговатыми, более заостренными на конце, выемка между лопастями листа сужается и углубляется (рис. 2).
Рисунок 2. Усредненные контуры листовых пластинок кувшинки для трех различных групп листьев, выделенных по их форме
| Nymphaea alba (рис. 1, «зона A») | N. candida (рис. 1, центральная область) | N. tetragona (рис. 1, зона «B») |
 |
 |
 |
Длина и ширина одной и той же листовой пластинки до и после гербаризации различается с высокой степенью достоверности (тест Стьюдента: N=32, t=2.9; Df=63; p=0.005 и t=3.9; Df=59; p=0.0003 соответственно). При гербаризации длина листовой пластинки без учета выбросов уменьшается в 1.05-1.4 раза (среднее арифметическое±стандартное отклонение 1.3±0.2) – эта величина отражает степень ее деформации в продольном направлении. Ширина листовой пластинки уменьшается в 1.15-1.5 раза (в среднем 1.35±0.2) – эта величина отражает степень ее деформации в поперечном направлении. Степень деформации листовой пластинки в продольном направлении достоверно не связана (r=0.3; p=0.1) со степенью ее деформации в поперечном направлении. Коэффициент деформации листовой пластинки (отношение степени ее деформации в продольном направлении к степени ее деформации в поперечном направлении) колеблется от 0.75 до 1.1 (в среднем 0,9±0,1).
Достоверные различия между размерами пыльцевых зерен одного и того же цветка до и после ацетолиза отсутствуют (тест Стьюдента: малый экваториальный диаметр: t=1.8, p=0.1; большой экваториальный диаметр: t=0.8, p=0.5, рис. 3).
Рисунок 3. Размеры пыльцевых зерен из гербаризованных цветков трех видов кувшинки (Nymphaea alba, N. candida, N. tetragona)
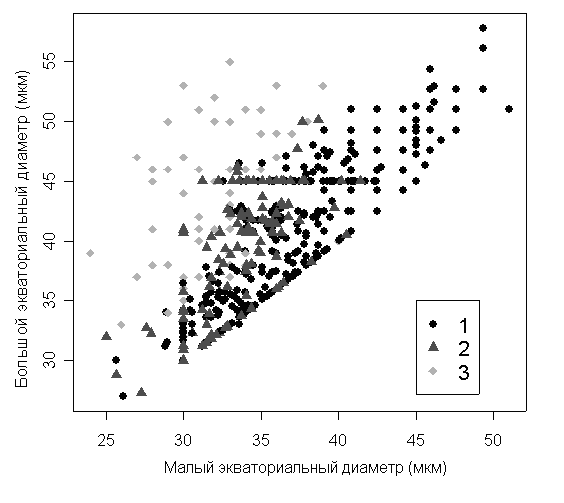
1 – отсутствие последующей обработки, 2 – ацетолиз, 3 – СЭМ.
Условия высокого вакуума, необходимого для СЭМ, приводят к заметной деформации пыльцевых зерен (рис. 3). Значения малого экваториального диаметра существенно уменьшаются при СЭМ (квартильный размах 30-36 мкм по сравнению с 37-46 мкм по данным промеров под световым микроскопом), эти различия высоко достоверны (тест Стьюдента: p<<0.01). Значения большого экваториального диаметра у пыльцевых зерен после СЭМ достоверно не изменяются (тест Стьюдента: p=0.56). Таким образом, на СЭМ фотографиях пыльцевые зерна оказываются гораздо более вытянутыми, чем они есть на самом деле (квартильный размах отношения большого экваториального диаметра к малому 1.0-1.1 до СЭМ и 1.2-1.5 после СЭМ), эти различия высоко достоверны (тест Стьюдента: p<<0.01).
Наше исследование показало, что даже при тщательной гербаризации непредсказуемым образом искажаются такие важные диагностические признаки, как размер и форма листовой пластинки кувшинки. Этот факт справедливо отмечался ранее (Uotila, 2001; Лисицына, 2003), хотя количественные исследования деформации листовых пластинок кувшинки при гербаризации на конкретных образцах были впервые проделаны лишь в настоящей работе. Надо отметить, что в связи со спецификой объекта качественная гербаризация кувшинок затруднена и требует больших временных затрат при сушке (Лисицына, 2003).
Нами также впервые показано, что характер различий формы листовой пластинки – часто используемого диагностического признака (Комаров, 1937; Маевский, 1976; Лисицына и др., 1993; Uotila, 2001) – до и после гербаризации чрезвычайно сходен с характером межвидовых различий (листья с формой листовой пластинки, типичной для N. alba, после гербаризации приобретают типичную для N. tetragona форму). В связи с этим нужно более критически относиться к систематическим обработкам рода, так как они в основном проводятся на основе гербарного материала (Комаров, 1937; Маевский, 1974; Лисицына и др., 1993; Определитель…, 1995), что вызывает дополнительные трудности в диагностике видов кувшинки. По этой причине чрезвычайно важно исследовать морфологию кувшинок на живом материале (Uotila, 2001).
Ацетолизная обработка не изменяет существенно размерные характеристики пыльцевых зерен. Поэтому данные, полученные на ацетолизированной пыльце, могут использоваться для сравнения с данными, полученными для необработанных пыльцевых зерен. Размерные характеристики пыльцы кувшинки существенно искажаются при СЭМ. В связи с этим измерения пыльцевых зерен кувшинки на СЭМ-микрографиях, сделанные J.B. Muntendam с соавторами (1996), нельзя использовать для сравнения с данными по необработанной или ацетолизированной пыльце.
Разумеется, полученные нами результаты не следует без проверки распространять на другие группы растений, учитывая в особенности своеобразную морфологию листьев (Heslop-Harrison, 1955) и пыльцевых зерен (Куприянова, 1976) кувшинки. Однако наши исследования подчеркивают необходимость количественного изучения характера изменений растительного материала при традиционных способах его фиксации в ходе таксономических исследований.
Искренне благодарна всем, кто оказывал всестороннее содействие при сборе материала в полевых условиях: участникам летних практик Московской Гимназии на Юго-Западе (№1543), проходивших под руководством С.М. Глаголева; зам. директора по НИР природно-исторического заповедника «Аркаим» Е. Чибилеву; зам. директора по НИР Кандалакшского заповедника А.С. Корякину, доценту Липецкого университета К.И. Александровой, директору Байкальского заповедника В.И. Сутула и зам. директора по НИР Астраханского заповедника А.К. Горбунову. Благодарна также сотруднице каф. высших растений биологического факультета МГУ С.В. Полевовой за консультации в области палинологии, сотруднику зоологического музея МГУ И.Я. Павлинову за консультации в области геометрической морфометрии, А.Б. Шипунову и С.Р. Майорову за конструктивное обсуждение работы и ведущему инженеру МЛЭМ А.Г. Богданову за компетентную и доброжелательную помощь на сеансах электронной микроскопии.
Комаров В.Л. Кувшинка – Nymphaea L. // Флора СССР. Л.; М, 1937. Т. 7. С. 6—12.
Крупкина Л.И. Nymphaeceae Salisb. — Кувшинковые // Флора Восточной Европы. СПб, 2001. Т. 10. С. 25—30.
Куприянова Л.А. Морфология пыльцы видов Nymphaea европейской части СССР // Бот. журн. 1976. Т. 61. N 11. С. 1558—1563.
Лисицына Л.И. Гербаризация водных растений, оформление коллекций // Матер. школы по гидроботанике. Рыбинск, 2003. С. 49—56.
Лисицына Л.И., Папченков В.Г., Артеменко В.И. Род Nymphaea L. – Кувшинка // Флора водоемов Волжского бассейна. СПб, 1993. С. 136—137.
Маевский П.Ф. Флора средней полосы европейской части СССР. 9-ое изд. Л., 1964. 880 с.
Определитель сосудистых растений центра европейской части России / Ред. И.А. Губанов и др. 2-е изд., дополн. и перераб. М., 1995. 560 с.
Основы микротехнических исследований в ботанике: справочное руководство / Р.П. Барыкина, Т.Д. Веселова, А.Г. Девятов и др. М., 2000. 127 с.
Adams D.C., Rohlf F.J., Slice D.E. Geometric morphometrics: ten years of progress following the ‘revolution’ [electronic resource]. 2002. Mode of access: http://life.bio.sunysb.edu/morph/review/review.html.
Bookstein F.L. Morphometric tools for landmarks data. New York, 1991. 435 p.
Britton N., Brown A. An illustrated flora of Northern US, Canada and british possessions. 2nd ed. New York, 1913. 412 p.
Heslop-Harrisson J. Nymphaea // J. Ecol. 1955. Vol. 43. P. 719—734.
Muntendam J.B., Povel G.D.E., van der Velde G. Morphometric patterns in the Nymphaea alba-candida complex // Acta Bot. Neerl. 1996. Vol. 45. N 3. P. 279—302.
R Development Core Team. R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, 2004. ISBN 3-900051-00-3, http://www.R-project.org.
Rohlf F.J. tpsUtil – TPS utility program [Electronic resource]. 2000. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsRelw: relative warps. Version 1.23 [Electronic resource]. 2001a. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsDig. Version 1.23 [Electronic resource]. 2001b. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsSuper. Version 1.12 [Electronic resource]. 2003. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. tpsRegr. Version 1.31 [Electronic resource]. 2005. Mode of access: http://life.bio.sunysb.edu/morph/.
Vermeulen P. Studies in dactylorchids. Utrecht, 1947. 251 p.
Volkova P.A., Shipunov A.B. Morphological variation in genus Nymphaea L. in the European Russia // Ann. Bot. Fen. В печати.
Uotila P. Nymphaea L. // Flora Nordica. 2000. Vol. 2. P. 216—221.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |