Дафнии (род Daphnia O. F. Mueller, 1785) – планктонные организмы, относящиеся к надотряду ветвистоусых надкласса ракообразных. Внутри этого рода выделяется среди прочих группа близкородственных видов -- группа D. longispina. Она включает в себя такие широко распространённые на территории Европы виды, как D. hyalina, D. galeata, D. cucullata, D. longispina s. str. (Hebert et al., 1989, цит. по: Brancelj et al., 1996). Морфологические отличия между видами этой группы заключаются в основном в форме переднего края головы (выпуклая для D. galeata, вогнутая для D. longispina и прямая для D. hyalina) и форме рострума (длинный и острый для D. hyalina, короткий и затупленный для D. longispina и D. galeata).
В то же время известно, что изменчивость некоторых морфологических параметров у дафний, таких как длина хвостовой иглы или форма головы, зависит от условий обитания: присутствия в воде хищников (как позвоночных, так и беспозвоночных), концентрации растворённых в воде кайромонов (веществ, выделяемых хищниками), доступности пищи для дафний в водоёме. Так, например, появление рыб в водоёме вызывает уменьшение размеров тела и увеличение относительной длины иглы в популяциях дафний (Spaak, Boersma, 1997), а наличие хищных личинок Chaoborus flavicans вызывает изменение соотношений частот различных форм головы у особей среди различных размерных классов (Brancelj et al., 1996).
Таким образом, отмечаемая многими авторами (Brancelj et al., 1996; Spaak, Boersma, 1997) широкая фенотипическая изменчивость видов группы D. longispina может затруднять их разграничение по морфологическим признакам. Насколько нам известно, для видов этой группы изучение морфологической изменчивости на массовом материале до сих пор не было проведено, поэтому мы решили исследовать морфологическую изменчивость D. longispina на одном острове Белого моря.
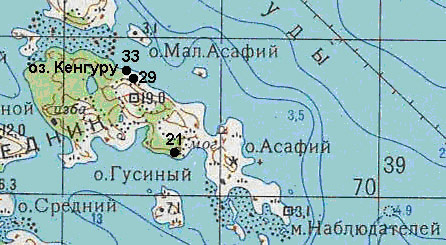
Половозрелые самки Daphnia longispina были собраны из пресноводных водоёмов на острове Асафий Кемь-Лудского архипелага Кандалакшского залива Белого моря. Одна из проб взята из озера «Кенгуру», три другие – из наскальных ванн (рис. 1, прил. 1). Сбор трёх проб из наскальных ванн проводился в последней декаде июля – первой декаде августа 2008 года. Проба из оз. Кенгуру была взята в конце июля 2007 года.
Так же, одновременно со сбором проб из водоёмов, мы проводили измерения параметров самих водоёмов, таких как линейные размеры, высота над уровнем моря, состав растительности, состав потенциальных хищников, питающихся дафниями, pH воды, а также отмечали наличие лишайников рода Xanthoria, как маркёра частых посещений птицами (прил. 2).
Мы описывали форму и размер тела дафний с помощью двух альтернативных подходов: классической морфометрии (совокупность линейных промеров тела) и геометрической морфометрии (метод, позволяющий оценить форму объекта как таковую, без учета его линейных размеров).
Рисунок 1. Схема расположения исследованных водоёмов на о. Асафий (Кемь-Лудский архипелаг, Кандалакшский залив Белого моря)
Точками обозначено расположение наскальных ванн (номера соответствуют использованным в тексте). Сторона квадрата километровой сетки соответствует 1 км на местности.
Рисунок 2. Схема промеров при использовании классической морфометрии и расположение меток при использовании геометрической морфометрии
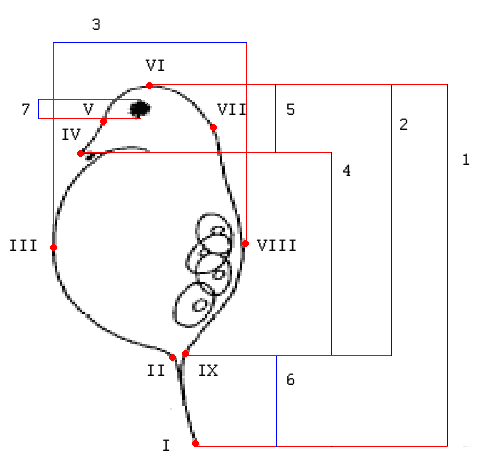
Арабскими цифрами обозначены морфологические промеры: 1 – общая длина тела (с иглой), 2 – длина тела (без иглы), 3 – ширина тела, 4 – длина карапакса, 5 – высота шлема, 6 – длина иглы, 7 – диаметр глаза. Римскими цифрами указаны порядковые номера меток, использованных при геометрической морфометрии (на рисунке – красные точки).
Рисунок 3. Распределение исследованных особей D. longispina на плоскости двух первых измерений анализа главных компонент матрицы морфологических промеров
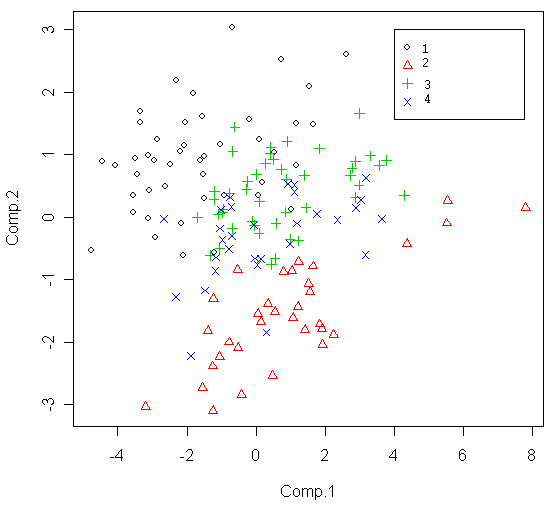
1 – образцы из скальной ванны №33, 2 – образцы из оз. «Кенгуру», 3 – образцы из скальной ванны №21, 4 – образцы из скальной ванны №29.
Рисунок 4. Распределение исследованных особей D. longispina на плоскости первых двух измерений анализа главных компонент матрицы частных деформаций
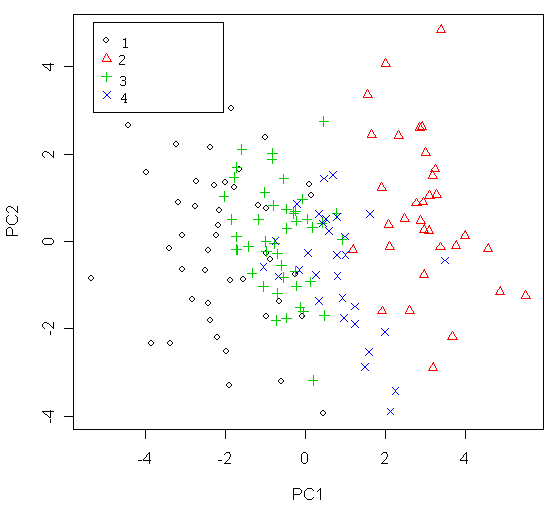
1 – образцы из скальной ванны №33, 2 – образцы из оз. «Кенгуру», 3 – образцы из скальной ванны №21, 4 – образцы из скальной ванны №29.
Рисунок 5. Половозрелые самки D. longispina из оз. «Кенгуру» (слева) и скальной ванны № 33 (справа). Хорошо видны различия по форме карапакса и относительной длины хвостовой иглы.
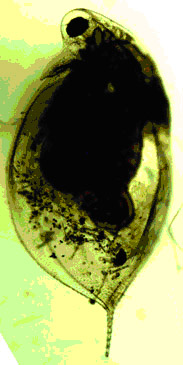 |
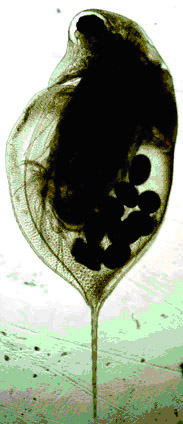 |
Brancelj A., Celhar T., Sisko M. 1996. Four different head shapes in Daphnia hyalina (Leydig) induced by the presence of larvae of Chaoborus flavicans (Meigen) // Hydrobiologia 339: 31–45.
Rohlf F.J. 2000. tpsUtil – TPS utility program. N.Y.: State Univ. at Stony Brook. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. 2001a. tpsRelw: relative warps. N.Y.: State Univ. at Stony Brook. Version 1.23. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Rohlf F.J. 2001b. tpsDig. N.Y.: State Univ. at Stony Brook. Version 1.23. [Electronic resource]. Mode of access: http://life.bio.sunysb.edu/morph/.
Spaak P., Boersma M. 1997. Tail spine length in the Daphnia galeata complex: costs and benefits of induction by fish // Aquatic Ecology 31: 89–98.
StatSoft, Inc. 1999. STATISTICA for Windows [Computer Program Manual]. Tulsa, OK.
Сбор материала проводился в рамках Беломорской экспедиции Московской гимназии на Юго-Западе (№1543) на территории Кандалакшского государственного природного заповедника. Мы признательны Е.В. Елисеевой, С.В. Сухову, Ю. Быкову и С. Иванову за помощь в сборе материала, а также П.Н. Петрову за большой вклад в обсуждение полученных результатов.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |