Ветвистоусые ракообразные (Cladocera) наскальных
луж островов Белого моря. Видовой состав и частоты встречаемости
Глаголев С., Быков Ю. Ветвистоусые ракообразные (Cladocera) наскальных
луж островов Белого моря. Видовой состав и частоты встречаемости // Материалы Беломорской экспедиции Московской Гимназии на Юго-Западе. Вып. 11 [Электронный ресурс]. 2011. Режим доступа:
http://herba.msu.ru/shipunov/belomor/2011/zoolog/luzhi.htm
Glagolev S., Bykov Yu. Cladocera in
rock pools on islands of the White sea. Species composition and frequency of
occurence // The materials of the White Sea Expedition
of Moscow South-West High School. Vol. 11 [Electronic resourse]. 2011. Mode of
access:
http://herba.msu.ru/shipunov/belomor/2011/zoolog/luzhi.htm [in Russian]
Введение
Наскальные лужи – углубления в скальном субстрате, хотя бы временно заполняющиеся водой. Они привлекают к себе всё большее внимание как модельные объекты для разнообразных экологических исследований [Srivastava
et al., 2004; Brendonck et al., 2010]. Каждая лужа – сравнительно простая экосистема с небольшим числом видов. Это позволяет на больших выборках водоемов изучать взаимосвязи между видами и факторы, влияющие на их распространение, динамику численности и генетический состав популяций.
Наскальные лужи распространены в разных ландшафтах по всему миру [Brendonck et al., 2010]. Лужи могут существенно отличаться друг от друга по солености, pH, первичной продукции и прочим параметрам среды. Они часто встречаются на побережьях водоемов и на островах. Лужи одного острова можно рассматривать как метасообщество, а населяющие их популяции образуют метапопуляции [Haag et al., 2005]. Совокупность луж на нескольких островах – еще более сложная система. Часто они расположены группами, а некоторые лужи могут образовывать каскады (во время дождя потоки воды стекают из одной лужи в другую). Кроме того, острова, на которых они находятся, отличаются по размерам и степени изолированности от материка и друг от друга.
Основу зоопланктона наскальных луж часто составляют ветвистоусые рачки (Cladocera). Наиболее детально факторы, влияющие на распределение видов Cladocera в наскальных лужах, были изучены на островах Балтийского моря [Ranta, 1979, 1982; Bengtsson 1986, 1988, 1989; Pajunen, Pajunen, 2003, 2007]. В этих работах показано, что на распределение видов дафний (Daphnia magna,
D. pulex, D. longispina) влияют такие факторы, как соленость, степень постоянства водоема (показатель, зависящий от глубины лужи) и межвидовая конкуренция. Однако распределение других видов Cladocera изучено в этих работах недостаточно подробно.
В Кандалакшском заливе Белого моря исследования зоопланктона наскальных луж одним из первых проводил А.М. Гиляров [Ghilarov, 1967]. Он исследовал 36 пресных и солоноватых луж на островах Кастьян, Наумиха, Асафий и Красный (два последних острова относятся к архипелагу Кемь-Луды) и отметил 9 видов ветвистоусых, определил проценты встречаемости и частоты совместной встречаемости. К сожалению, в работе не приведены данные о распределении изученных луж (и видов) по отдельным островам и о видовом составе отдельных луж. Фауна ракообразных наскальных луж Кандалакшского залива также была исследована в работе Н.В. Вехова [Вехов, 2001] путем однократного взятия проб на 23 островах, в том числе на о-вах Кемлудский, Воротиха и Красный архипелага Кемь-Луды и на о. Горелый Керетского архипелага. В пробах из 250 луж обнаружено 14 видов ветвистоусых, включая
два чисто морских – Podon intermedius Lilljeborg, 1853 (на самом деле, вероятно,
Podon leuckarti G.O. Sars, 1862 [Мордухай-Болтовской, Ривьер, 1987]) и
Evadne anonyx G.O. Sars, 1897 (на самом деле – Evadne nordmanni Lovén, 1836 [Мордухай-Болтовской, Ривьер, 1987]). Также фауну Cladocera на островах Кандалакшского залива изучали И.А. Cтогов и Н.В. Полякова с соавторами [Стогов и др., 1996; Полякова и др., 2006]. Для изученных островов Керетского архипелага (рис.1) ими указано 13 видов ветвистоусых.
Рисунок 1. Район исследований
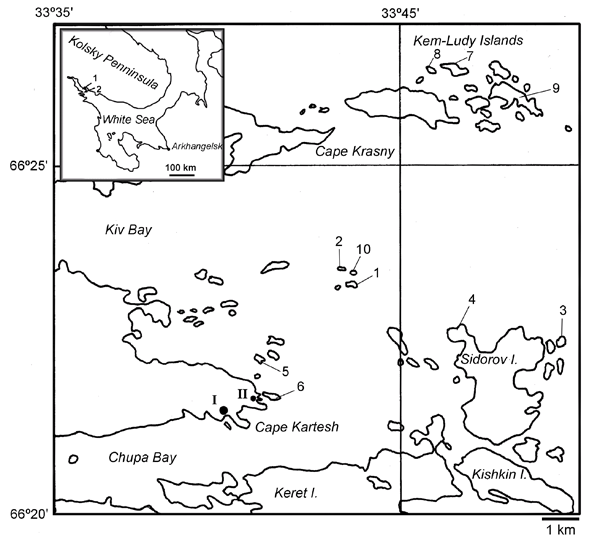
На врезке: 1 – острова Средние Луды, 2 – расположение района исследований. Исследованные острова: 1 – о. Большая Одинокая Луда (66˚22’25’’ с.ш., 33˚44’01’’ в.д, 7 луж), 2 – о. Средняя Одинокая луда (66˚22’39’’ с.ш., 33˚43’49’’ в.д., 7 луж), 3 – о. «Черепаха» (66˚21’27’’ с.ш., 33˚52’24’’ в.д., 7 луж), 4 - о. Сидоров (66˚21’41’’ с.ш., 33˚48’18’’ в.д., 4 лужи), 5 – о. «Гроздовник» (66˚21’10’’ с.ш., 33˚40’25’’ в.д., 2 лужи), 6 – о. Иваньков (массовое обследование, см. текст), 7 – о. Коржничиха (66˚25’57’’ с.ш., 33˚47’43’’ в.д., 1 лужа), 8 – о. «Кораблик» (66˚25’52’’ с.ш., 33˚47’01’’ в.д., 3 лужи), 9 – о. Асафий (66˚25’22’’ с.ш., 33˚50’09’’ в.д., 23 лужи), 10 – о. Малая Одинокая луда (массовое обследование, см. текст). I – Биостанция ЗИН РАН, II – базовый лагерь экспедиции. В кавычках даны условные названия, присвоенные безымянным островам во время беломорской экспедиции Московской гимназии № 1543.
Гиляров (1967) и Вехов (2001) использовали в своих работах метод однократного взятия проб. Стогов (1996) и Полякова (2006) с соавторами проводили долговременные наблюдения, но в их работах не содержится данных по отдельным островам и группам островов. Мы решили провести долговременные исследования состава и динамики фауны ветвистоусых в наскальных лужах островов Белого моря.
В 2001 г. мы начали изучать наскальные лужи на островах Керетского и Кемь-Лудского архипелагов, а также некоторых островах губы Кив (Кандалакшский залив Белого моря). Целью работы было исследование состава фауны ветвистоусых, а также выявление закономерностей распределения видов ветвистоусых на разных островах и в разных лужах в зависимости от их параметров. Предварительные данные этих исследований за 2003-2005 годы были опубликованы нами ранее [Глаголев и др., 2007]. В данном сообщении приводятся результаты многолетних наблюдений за видовым составом и частотами встречаемости ветвистоусых в изучаемых водоемах.
Материалы и методы
Многолетние исследования
В 2003-2005, а также в 2007 и 2008 годах по стандартной методике были исследованы 54 лужи на 8 островах в районе губы Чупа в Кандалакшском заливе Белого моря (рис.1). Эти острова образуют три группы: 1) острова архипелага Кемь-Луды (Асафий,
«Кораблик»,
Коржничиха); 2) острова Кив-губы (Большая и
Средняя Одинокая Луды,
«Гроздовник»); 3) острова Керетского архипелага (Сидоров,
«Черепаха»). Большинство луж обследовали один раз в год в конце июля – начале августа. В 2007 и 2008 годах лужи на островах «Гроздовник», Большая Одинокая Луда, Средняя Одинокая луда, «Черепаха», Сидоров (всего 30 луж) обследовали повторно спустя 17 дней, чтобы установить, насколько сильно меняется видовой состав ветвистоусых в одной луже в течение короткого периода.
У выбранной лужи измеряли максимальную длину и ширину рулеткой с точностью до 5 см, а максимальную глубину линейкой с точностью до 1 см. Площадь нескольких первых луж измеряли «по клеточкам» с помощью куска полиэтилена, расчерченного на квадратные метры и дециметры, а у остальных луж – на глаз; сравнение этих данных показало, что при визуальной оценке площади луж результат не более чем в 1,5 раза отличается от полученного при измерении «по клеточкам». Высоту лужи над уровнем моря определяли также на глаз. Приняв форму лужи за конус, вычисляли ее объем как площадь, умноженную на 1/3 максимальной глубины. Кислотность воды в луже измеряли с помощью портативного рН-метра фирмы Hanna Instruments (точность до 0,1 единицы рН), солёность – портативным кондуктометром ExStick EC400 (EXTECH instruments).
Рядом с каждой лужей на камнях ставили метку несмываемой краской, что позволяло обнаружить ее в последующие годы. Отмечали наличие на камнях рядом с лужей (на расстоянии до 1 м) лишайника Xanthoria как показатель частого посещения данной точки птицами. Этот лишайник нуждается в больших количествах соединений азота, содержащихся в птичьем помете [Armstrong, 1984]. В луже отмечали наличие других таксонов животных – в первую очередь, потенциальных хищников.
Из луж планктонным сачком (газ № 68) брали пробы и фиксировали 96% этанолом. Определение видов в большинстве случаев проводили по живому материалу в полевых условиях с использованием микроскопа. В сложных случаях мы обращались за помощью к специалистам по систематике ветвистоусых (см. Благодарности).
Массовое обследование луж
В 2007, 2008 и 2010 годах обследовали большее число луж на некоторых островах, чтобы выяснить, насколько сильно определенные нами по небольшой выборке луж в многолетних исследованиях частоты встречаемости видов отличаются от частот, выявленных на тех же островах по большей выборке. В 2007 году обследовали острова Большая Одинокая Луда (40 луж), Средняя Одинокая Луда (23 лужи), «Черепаха» (34 лужи), Асафий (28, отличные от регулярно обследуемых). В 2008 году обследовали только остров Малая Одинокая Луда (17 луж), в 2010 году – о. Большая Одинокая Луда (46 луж), Средняя Одинокая Луда (45 луж) и «Черепаха» (45 луж). В 2007 г. отмечали только виды ветвистоусых в лужах, в 2008 и 2010 годах – еще и морфометрические параметры луж. В 2010 г. обследовали также 59 луж на о. Иваньков (Кив-губа, см. рис. 1). Ранее, в 2001-2005 г.г., на этом острове обследовали 6 луж. В 2007 и 2010 году на островах было подсчитано общее число луж. При этом лужей считали водоем с соленостью менее 6 ‰, глубже 10 см и диаметром более 1 метра. На крупном о. Асафий подсчитывали лужи только в прибрежной зоне не далее чем в 100 м от берега.
В 2011 г. 1 августа обследовали 23 лужи на наиболее удаленных от материка островах Средние Луды (13 на
большом острове и 10 на двух соседних мелких островках) и 12 луж на
о. Наумиха.
Обработку данных проводили в статистической среде R 2.9.0 (R Development Core Team, 2004). Достоверность различия частот встречаемости видов между парами островов проверяли при помощи теста пропорций. Построение кривых накопления проводили при помощи программы EstimateS 8.2.0 [Colwell, 2006] с использованием функции построения Sample-based Rarefaction curve [Colwell et al. 2004].
Результаты
Общее число луж на острове по данным 2007 года составило от 10 (о. «Кораблик», о. «Гроздовник») до 260 (о. Асафий). На Средней Одинокой Луде найдено 50 луж, на Большой Одинокой Луде 75, на о. «Черепаха» около 150 луж. В 2010 году число луж на Средней и Большой Одинокой Лудах подсчитывали после длительного периода сухой погоды. Найдено по 45 луж. На о. «Черепаха» после периода дождей в 2010 году найдено 324 лужи, а на о. Иваньков около 60. На о. «Черепаха» 221 лужа располагалась на мористой стороне, а на стороне, обращенной к о. Сидоров, луж было в 2 раза меньше (113). По нашим наблюдениям, такое распределение луж характерно и для других островов. Большая часть луж расположена на их мористой, более пологой стороне. Число луж слабо коррелирует с размером острова: на островах сходного размера (например, «Черепаха», Иваньков, Большая Одинокая Луда) оно может резко различаться.
Площадь выбранных луж варьировала от 0,3 до 50 м2, средняя площадь 11 м2, глубина – от 10 до 150 см, средняя глубина 34 см (по данным 2003-2007 гг).
Видовой состав
В течение трех лет на обследованных островах стабильно обнаруживали 12 видов ветвистоусых (табл. 1).
Из эпизодически встречающихся видов в пробах 2001 года с острова Сидоров найден вид
Alonella excisa, который за последующие годы не был найден ни в одной луже (правда, данная лужа в дальнейшем не была обследована, а на о. Асафий единожды отмечен не определенный нами вид Chydoridae, отличный от
C. sphaericus). Сeriodaphnia pulchella была обнаружена в пробах 2003 г. с двух островов – «Черепаха» и Сидоров. В дальнейшем из-за сложности определения в полевых условиях оба вида рода
Ceriodaphnia учитывались в нашей работе как Ceriodaphnia sp. Судя по обработанным пробам, в подавляющем большинстве случаев это была
C. quadrangula.
Из девяти «массовых» видов (средняя частота встречаемости за все годы превышает 5%) нет ни одного, который не встретился бы хотя бы один раз за все годы на каждой из групп островов. Однако некоторые «массовые» виды встречаются на отдельных островах лишь эпизодически. Например, на Одиноких Лудах
D. longispina была отмечена единственный раз за все время наблюдений, причем только на Большой Одинокой Луде в единственной луже.
P. pediculus был отмечен на этих островах только дважды: в 2008 и 2010 гг., и только на Средней Одинокой Луде.
B. longirostris была обнаружена на Одиноких Лудах только в 2003 году (на Большой Одинокой Луде) и в 2010 году (на Средней Одинокой Луде).
D. curvirostris не была найдена на Одиноких Лудах в первые три года наблюдений, но в последующие годы (2007, 2008, 2010) обнаруживалась на обоих островах. При массовом обследовании луж в 2010 году
D. curvirostris была найдена в том числе и в тех лужах, в которых этот вид ранее отсутствовал.
На некоторых островах отмечено не только эпизодическое появление, но и эпизодическое «исчезновение» отдельных видов. Например, на о. «Черепаха» некоторые виды не были найдены только в отдельные годы. В 2010 году при наиболее массовом обследовании луж не были найдены такие регулярно встречающиеся на этом острове виды, как
P.pediculus, D. curvirostris и D. pulex.
На о.Асафий из года в год видовой состав ветвистоусых не менялся.
На о. Иваньков во все годы наблюдений обнаружены только 3 вида ветвистоусых:
D. magna, C. sphaericus, S. mucronata.
В 2011 году при однократном посещении на Средних лудах и на о. Наумиха обнаружено по 5 видов ветвистоусых:
D. magna, D. curvirostris, C. sphaericus, S. mucronata и
Ceriodaphnia sp.
Частоты встречаемости
Для каждого «массового» вида вычислили частоту его встречаемости в районе исследований за все годы (табл. 1). Наибольшую частоту встречаемости (выше 50%) имеют
D. magna и C. sphaericus. Частоту встречаемости 20-30 % имеют Ceriodaphnia
sp. и S. mucronata. Ниже (10-20%) частота встречаемости D. pulex,
B. longirostris, D. curvirostris. Наиболее низкую частоту встречаемости (менее 10%) имеют
D. longispina и P. pediculus. Без учета массовых обследований луж несколько увеличивается частота встречаемости
P. pediculus. Это связано с тем, что наибольшее количество луж при массовом обследовании было изучено на Одиноких Лудах, где данный вид редок.
В отдельные годы средние частоты встречаемости видов в районе исследований варьировали (табл. 2). У большинства видов они различались не более чем в 1.5-2 раза. Например, у
C. sphaericus от 48 до 70 % при средней частоте 58 %, у S. mucronata от 18 % до 39 % при средней частоте 29 %, а у
D. curvirostris от 4% до 9% при средней за все годы 7%. У P. pediculus частота встречаемости варьировала от 7 до 23% (средняя за все годы 15% без учета массовых обследований).
По данным ежегодных обследований 54 луж на 8 островах в 2003-2005, 2007 и 2008 годах и массовых обследований 278 луж на 5 островах в 2007 и 2010 гг. были вычислены частоты встречаемости видов ветвистоусых на разных островах и группах островов (табл. 2, 3).
Сравнение частот встречаемости видов на различных островах представлено в
табл. 4. Вопреки ранее опубликованным данным по результатам обследования маркированных луж [Глаголев и др., 2007], при массовых обследованиях выявлены достоверные различия частот встречаемости на разных островах.
Группы луж
На острове Асафий (Кемь-Луды) ежегодно обследовали 33 лужи. По расположению на острове их можно разбить на 7 групп (рис. 2). Расстояние между группами в большинстве случаев превышало 200 м, а расстояние между двумя соседними лужами в одной группе не превышало 30 м. Средняя встречаемость видов за все годы в группах луж на острове Асафий представлена в
таблице 5.
Рисунок 2. Группы луж на о. Асафий (архипелаг Кемь-Луды). Серым показано внутреннее озеро

Из данных таблицы видно, что лишь в двух группах из шести за 5 лет наблюдений обнаружены все виды, представленные на острове. В остальных четырех группах за все годы наблюдения отсутствовали некоторые виды ветвистоусых. Например, в первой группе из 6 луж ни разу не встречен такой часто встречающийся вид, как
D. magna, а также D. pulex. Во второй группе, также состоящей из 6 луж, не встречены
B. longirostris, P. pediculus и Ceriodaphnia sp. При этом в каждый отдельный год ни в одной группе луж не встретились все виды ветвистоусых.
С другой стороны, в каждый год при обследовании всех луж обнаруживались все виды с единственным исключением – в 2004 году на о. Асафий не найдена
D. pulex.
Нами для о. Асафий и о. Большая Одинокая Луда были построены «кривые накопления», показывающие как число обнаруженных на острове видов должно зависеть от числа взятых проб [Gotelli&Colwell, 2001]. Судя по этим кривым, полученным при наиболее массовом обследовании луж (рис. 3), для полного выявления состава фауны ветвистоусых на острове необходимо взять пробы из 25-50 луж. Те же выводы подтверждаются данными по о. «Черепаха», где
D. magna была обнаружена только в одной луже из 7 ежегодно обследуемых, в то время как после обследования большего числа луж выяснилось, что частота её встречаемости на острове в 3 раза выше.
Рисунок 3. Зависимость количества видов на острове от количества обследованных луж (по данным 2007 г)
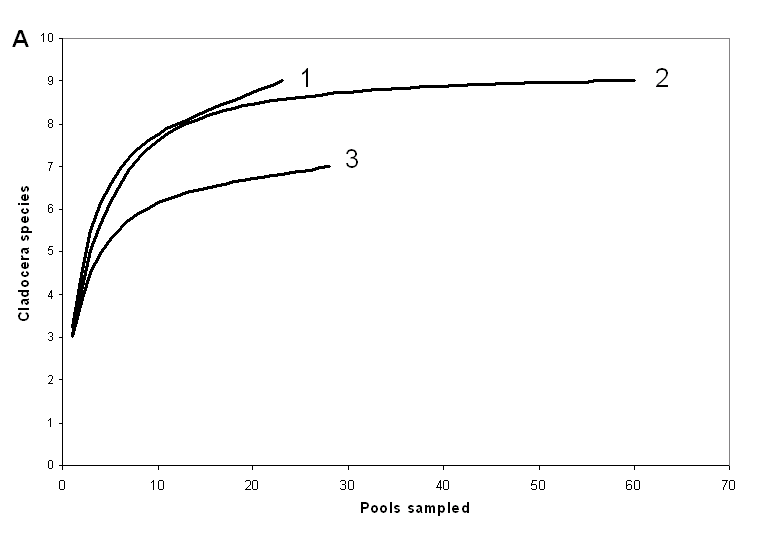
А – о. Асафий. 1 – ежегодно обследуемые лужи, включаемые в выборку, 2 – ежегодно обследуемые лужи + дополнительно обследованные, 3 – дополнительно обследованные лужи.
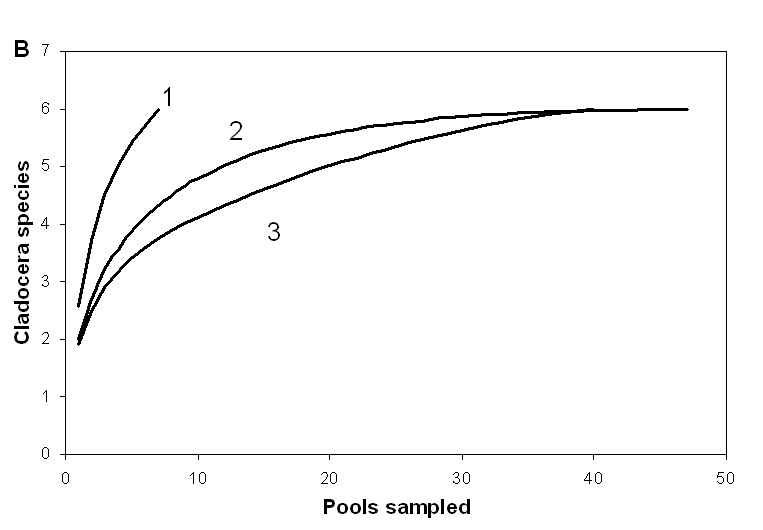
B – о. Большая Одинокая Луда. 1 – ежегодно обследуемые лужи, 2 – ежегодно обследуемые лужи + дополнительно обследованные, 3 – дополнительно обследованные лужи.
Обсуждение
Видовой состав
Нами впервые для островов Белого моря отмечены A. excisa и D. curvirostris. Последний вид, вероятно, отмечен впервые для всего беломорского региона, хотя он хорошо отличается от всех остальных видов европейских дафний (рис. 4) и весьма обычен для наскальных луж. Возможное неверное определение его как
D. pulex могло приводить к искажениям результатов экологических исследований.
Рисунок 4. Морфологические отличия D. pulex и D. curvirostris

А – рострум D. pulex, основание антеннул (отмечено стрелкой) не редуцировано, Б – рострум D. curvirostris, основание антеннул (отмечено стрелкой) редуцировано.
A. curvirostris все годы обнаруживался из сотен обследованных нами луж лишь в единственной, расположенной на вершине острова «Черепаха» и больше напоминающей водоем на сфагновом болоте, – типичное местообитание данного вида [Fryer, 1980]. Поэтому указание данного вида как типичного для наскальных луж [Полякова и др., 2006], на наш взгляд, вряд ли оправдано.
К редким для наскальных луж видам относятся также A. excisа и M. hirsuticornis. Оба вида были обнаружены на самом крупном из исследованных островов – на о. Сидоров, условия на котором наиболее близки к материковым. Однако
M. hirsuticornis также был единожды обнаружен на небольшом безлесном острове (о. «Кораблик»), что говорит о его способности расселяться на малые острова и выживать в существующих там условиях.
В качестве обычного для наскальных луж вида Стогов и соавторы [Стогов и др., 1996] указывают
Alonella nana (Baird, 1850). Нами этот вид на обследованных островах Керетского архипелага не найден.
В работах А.М. Гилярова [Ghilarov, 1967] и Н.В. Поляковой с соавторами [Полякова и др., 2006] в том же районе были отмечены другие виды:
Bosmina obtusirostris G.O. Sars, 1862, Daphnia arctica Werestschagin, 1913 и
Ceriodaphnia affinis Lilljeborg, 1900. Однако наличие этих видов в лужах не подтверждается.
Daphnia arctica, вероятно, не представляет собой самостоятельного вида и должна быть сведена в синонимы
Daphnia lacustris G.O. Sars, 1862 [С.М. Глаголев, неопубликованные данные, Nilssen et al., 2007].
Daphnia lacustris также не обнаружена нами, однако на исследованных островах встречаются похожие на этот вид морфотипы
D. longispina [Царев и др., 2011]. Ревизия рода Bosmina [Lieder, 1983, Sacherov, Havel, 1997] существенно изменила представления о границах видов и об их диагностических признаках. Ревизия рода
Ceriodaphnia не проведена, и точное определение многих видов затруднено.
Для наскальных луж Стогов и соавторы [Стогов и др., 1996] указывают также Daphnia cristata G.O. Sars, 1862,
Diaphanosoma brachyurum (Lievin, 1848) и Holopedium gibberum Zaddach, 1855, которые нами не обнаружены. Хотя ими были обследованы другие острова Керетского архипелага, вряд ли состав фауны на этих островах может столь радикально отличаться. Более того,
D. cristata и H. gibberum – типичные озерные виды, практически никогда не встречающиеся во временных водоемах. Их наличие в наскальных лужах требует подтверждения. Однако данные, представленные в более позднем сообщении Стогова и соавторов (2010), лучше совпадают с нашими.
Еще один вид, отмеченный на других островах Кандалакшского залива в работе Н.В.Вехова [Вехов, 2001], а нами не найденный
— Simocephalus vetulus (O.F.Müller, 1776). Он обнаружен Веховым на двух островах в кутовой части Кандалакшского залива и на о. Кастьян – вне района наших исследований.
Ceriodaphnia rotunda G.O. Sars, 1862 обнаружена Веховым на пяти островах, в том числе двух островах Кемь-Лудского архипелага, не обследованных нами. Вид указан как редкий (частота встречаемости меньше 5%). Возможно, он изредка встречается в наскальных лужах, но нами не найден. В то же время на Кемь-Лудах Веховым не был отмечен
P. pediculus (довольно обычный там вид).
Мы не обнаружили различий в составе фауны на разных группах островов, на существование которых указывают более ранние работы [Вехов, 2001]. Видимо, расстояние между островами в 2-3 км не является серьезной преградой для распространения Cladocera, а на каждом острове имеются более или менее подходящие местообитания для всех этих видов. На Средних Лудах, наиболее изолированной группе островов, и на о. Наумиха мы обнаружили по 5 видов ветвистоусых, хотя в работе Вехова [Вехов, 2001] указано лишь 3 вида для Средних Луд, но 8 видов для о. Наумиха. На о. Наумиха мы обследовали всего 12 луж, поэтому мы не можем уверенно судить о фауне ветвистоусых на данном острове. Не исключено, что, как и при обследовании Одиноких Луд, многократное посещение Средних Луд и о. Наумиха с исследованием большего количества луж позволило бы выявить на этих островах и другие виды ветвистоусых. Однако отсутствие на обследованных нами островах
S. vetulus явно показывает, что некоторые качественные региональные отличия фауны ветвистоусых на островах всё-таки существуют.
Поскольку D. longispina была найдена на о. Большая Одинокая Луда только один раз (в 2003 г.) в одной луже, а в последующие годы не обнаружена ни разу даже при обследовании большого количества луж, создается впечатление, что на этом острове она действительно исчезла. Возможно,
B. longirostris появляется на Одиноких Лудах лишь эпизодически или же присутствует в очень небольшом проценте луж, где мы не смогли ее обнаружить с 2004 по 2010 год.
P. pediculus, возможно, вселился на эти острова в 2008 году и закрепился на них до 2010 года. Похожая ситуация наблюдается и в случае
D. curvirostris. Чтобы подтвердить, что некоторые виды действительно появляются на одних островах и исчезают на других, необходимо тотальное обследование луж в течение нескольких лет.
Частоты встречаемости
К сожалению, в работе Вехова [Вехов, 2001] не приводятся частоты встречаемости видов на отдельных островах или группах островов, поэтому его данные трудно сравнить с нашими. Тем не менее, к редким видам в работе Вехова отнесены
C. pulchella и M. hirsuticornis, а к фоновым — D. magna и
C. sphaericus. В этом его данные совпадают с нашими. Но в качестве третьего фонового вида в работе Вехова указана
D. pulex (найдена во всех обследованных точках, частота встречаемости 10-80%). Даже если учесть, что Вехов не разграничивал
D. pulex и D. curvirostris [Н.В. Вехов, личное сообщение], по нашим данным суммарная частота встречаемости этих видов не достигала 80% ни в один год ни на одном из обследованных островов и, как правило, не превышала 30-40%.
По числу видов, имеющих достоверно разную частоту встречаемости, наиболее сильно отличаются друг от друга такие пары островов, как Большая и Средняя Одинокие Луды, Средняя Одинокая Луда и Асафий, Большая Одинокая Луда и «Черепаха». Судя по этим данным, степень различий не связана ни с размерами островов, ни с расстоянием между ними.
Наиболее обеднен видовой состав ветвистоусых на о. Иваньков, который в отлив соединяется с материком, но, тем не менее, не удален на большое расстояние от других, более мористых островов. На нем присутствуют всего 3 вида, причем
D. magna при массовом обследовании 2010 г. найдена лишь в одной луже из 37. По данным В.Н.Вехова [Вехов, 2001], столь же сильная обедненность характерна для материкового побережья, обследованного им в четырех пунктах, где им отмечены всего 4 вида пресноводных ветвистоусых:
D. magna, D. pulex, C. sphaericus и B. longirostris. Поэтому можно предполагать, что обедненный видовой состав характерен для материковых наскальных луж.
По площади остров Иваньков сравним с такими островами, как «Черепаха» и Большая Одинокая луда. Число луж на острове, вероятно, лишь ненамного меньше, чем на Большой и Средней Одинокой лудах. Чем же объясняется обедненность его фауны? Хотя этот остров был обследован в 2010 году после продолжительной засухи и последовавших за ней сильных дождей, отсутствие ветвистоусых примерно в половине луж не было связано с их пересыханием: во многих лужах присутствовали взрослые личинки Chironomidae, так что эти лужи полностью не пересыхали. Всё же пересыхание луж может играть некоторую роль. Даже после периода дождей практически все лужи на о. Иваньков имели глубину менее 30 см, в то время как на других островах встречались и более глубокие лужи. Можно предположить, что средняя частота высыхания луж на этом острове больше. Показано [Altermatt et al., 2009], что предсказанная по морфометрическим параметрам частота пересыхания луж отрицательно коррелирует с ежегодным наличием в них трех видов
Daphnia. Возможно, частое пересыхание луж — один из факторов, обуславливающих обедненный видовой состав о. Иваньков. Однако один этот фактор не может объяснить отсутствие многих видов, так как в лужах той же глубины на других островах их частота встречаемости была достаточно высокой.
Некоторое влияние на видовой состав может оказывать и общее число луж на острове. Число луж на о. Иваньков значительно меньше, чем на сравнимом с ним по размерам мористом о. Черепаха, где в этот же период насчитывалось в 5 раз больше луж, и сравнимо с таковым на о. Средняя Одинокая Луда. Видовой состав ветвистоусых наиболее богат на о. «Черепаха», на о. Средняя Одинокая Луда несколько обеднен и на о. Иваньков наиболее беден (табл. 3). При увеличении среднего расстояния между лужами уменьшается обмен видами ветвистоусых и повышается вероятность вымирания видов; ранее предполагалось, что этот фактор может влиять на обедненность видового состава водных жуков в наскальных лужах [Ranta, 1982].
В близлежащих материковых водоемах обитают такие виды, как D. longispina,
P. pediculus, S. mucronata, B. longirostris, C. sphaericus [Куликова, 2001], но, вероятно, отсутствуют остальные массовые виды островных луж. Тем не менее, на о. Иваньков отсутствуют как некоторые «материковые», так и практически все «островные» виды.
Можно предположить, что этот остров гораздо меньше посещается птицами (в отлив он соединяется с материком, и морские птицы на нем, видимо, почти не гнездятся). Признак редкого посещения птицами о. Иваньков
— практически полное отсутствие на нем лишайника Xanthoria, который предпочитает скалы с большим количеством птичьего помета. Возможно, это уменьшает частоту попадания эфиппиумов ветвистоусых с других островов и из материковых водоемов. Однако относительная роль птиц, ветра и воды в расселении ветвистоусых на островах не изучена; возможно, ветер играет более важную роль, чем птицы [Bohonak, Jenkins, 2003].
Птицы могут оказывать и иное влияние на распределение ветвистоусых. Оставляя на островах помет, они обогащают почву и водоемы, вызывая эвтрофикацию луж и увеличивая первичную продукцию. Даже в крупных озерах птичий помет может служить основным источником фосфора, содержание которого часто лимитирует первичную продукцию [Manny et. al., 1994]. Известно, что
D. magna предпочитает подобные богатые биогенными элементами местообитания [Koivisto, 1995]. Кроме того, увеличение количества пищи может способствовать снижению конкуренции и увеличению видового разнообразия ветвистоусых. Наконец, в обогащенных биогенами лужах ветвистоусые могут достигать большей численности и оставлять большее количество покоящихся яиц, что может способствовать выживанию и более активному расселению. Возможно, из определяющих обедненность видового состава «материковых» луж факторов решающее значение имеет их меньшее разнообразие: малая глубина, отсутствие луж, обогащенных биогенными элементами и др. Для проверки этой гипотезы необходимы дополнительные исследования, в том числе полевые эксперименты по обогащению луж биогенами и вселению в них ветвистоусых.
Наиболее удивительная и мало обсуждаемая в литературе особенность фауны ветвистоусых в наскальных лужах – почти полное отсутствие в них видов семейства Chydoridae. G. Fryer [Fryer, 1985] отмечает, что от размеров водоема наиболее сильно зависит именно число видов хидорид: в мелких водоемах их намного меньше. В этой же работе отмечается, что причины этого неизвестны. Предполагается, что данный эффект может быть связан с меньшим разнообразием водной растительности и с особенностями откладки эфиппиумов хидоридами (многие виды приклеивают их к растениям). Однако до сих пор эти причины не ясны; этот интересный вопрос явно требует дополнительного изучения.
Группы луж
Наши данные показывают, что даже многолетнее обследование произвольно взятой группы из 5-6 луж не дает полного представления о видовом составе ветвистоусых на таком острове, как о.Асафий. В то же время обследование 30 луж позволяло выявить все виды ветвистоусых даже при однократном взятии проб почти во всех случаях. Можно сравнить исследование фауны наскальных луж на острове с исследованием фауны озер. Показано, что для полного охвата фауны зоопланктона одного озера необходимо взять около 25-50 проб в различных биотопах [Dumont & Segers, 1996]. При исследовании фауны луж на острове также необходимо исследовать 25-50 разнообразных водоемов, представляющих собой различные биотопы островного метасообщества. Поэтому данные Вехова [Вехов, 2001], исследовавшего в среднем около 10 луж на каждом острове или участке побережья, возможно, не отражают в полной мере ни видового состава, ни реальных частот встречаемости видов на разных островах.
Из наших данных и данных других авторов следует, что ни видовое разнообразие ветвистоусых на отдельных островах, ни степень различия видового состава между отдельными островами на изученной выборке островов не зависят от их размеров, удаленности друг от друга и от материка так, как предсказывают классические представления равновесной теории островной биогеографии [MacArthur, Wilson, 1967]. Наиболее ярко об этом свидетельствует обедненность видового состава материковых луж и луж околоматериковых островов. Скорее на число видов ветвистоусых влияет наличие на острове луж, отличающихся друг от друга по глубине и степени постоянства, степени эвтрофикации, рН и солености, видовому составу других беспозвоночных, в том числе хищников, и т. д. Мы предполагаем, что равновесные модели могут не соответствовать наблюдаемому видовому разнообразию прежде всего из-за высокой расселительной способности ветвистоусых, которая позволяет многим видам с высокой частотой заселять любые острова вне зависимости от их размеров и удаленности.
Заключение
На островах встречается 3 группы видов ветвистоусых:
- A. excisa, M. hirsuticornis, A. curvirostris – на острова заносятся редко и для островной фауны не типичны, а обитают в озерных и болотных материковых водоемах;
- виды, которые встречаются как в материковых водоемах, так и в наскальных лужах островов –
P. pediculus, D. longispina, B. longirostris, S. mucronata,
C. sphaericus;
- виды, характерные только для наскальных луж – D. magna, D. curvirostris,
D. pulex, C. quadrangula.
Видовой состав рода Ceriodaphnia в наскальных лужах островов Белого моря требует дополнительного изучения.
В предыдущих исследованиях упускалось наличие D. curvirostris, имеющей некоторое внешнее сходство с
D. pulex, что могло приводить к искажению фаунистических и экологических данных. Наличие этих двух легко различимых видов [Глаголев, 1995] необходимо учитывать всем, кто будет в дальнейшем исследовать экологию зоопланктона наскальных луж.
Для полного выявления фауны ветвистоусых на острове необходимо взять пробы не менее чем из 25-50 луж.
Благодарности
Работа выполнена на базе ежегодной Беломорской экспедиции Московской гимназии на Юго-Западе № 1543. Определение видов
Ceriodaphnia было подтверждено Н.М. Коровчинским, вида Bosmina – А.В. Котовым, видов Chydoridae – Н.Н.Смирновым, за что авторы им глубоко признательны. Часть материала собрана на территории Кандалакшского государственного природного заповедника. Авторы выражают глубокую благодарность дирекции заповедника и в особенности А.С. Корякину, а также участникам экспедиции за содействие в сборе материала. Отдельно нам хотелось бы поблагодарить
А.Н. Квашенко, В.Я. Бергера,
Н.П. Дацкевича,
С.В. Сухова за техническую поддержку,
Е.В. Елисееву,
П.Н. Петрова,
П.Н. Дацкевича,
Д. Бережного,
И.В. Федорова, О.А. Злобовскую,
С.А. Касацкую,
В.А. Чаву, И.С. Юфрякова,
С. Иванова, Д. Закрасину,
П.Б. Борисову, М.С. Бизина,
И.М. Флямера, А. Кириченко за их помощь в сборе и обработке материала,
П.А.Волкову, E. П. Альтшулера и
А.Б. Шипунова за помощь в статистической обработке данных и ценные советы. Исследования частично финансировались грантом № P11-018 фонда «Династия»
и Московским институтом открытого образования.
Литература
Вехов В.Н., Ракообразные наскальных ванн островов и побережья Кандалакшского залива Белого моря. // Биология внутренних вод, 2001, № 3, с. 20-28.
Глаголев С.М., Род Daphnia // Определитель пресноводных беспозвоночных России и сопредельных территорий(под. ред. С.Я.Цалолихина), т.2. Ракоообразные, СПб, 1995, с . 48-58. Key to freshwater invertebrates of Russia and adjacent lands (ed. by S.J.Tsalolikhin)
Глаголев С.М., Дацкевич П.Н., Злобовская О.А., Чава В.А. 2007. Видовой состав и закономерности распределения ветвистоусых ракообразных (Cladocera) в наскальных лужах островов Белого моря. // Экологические исследования беломорских организмов (под ред. В.Я.Бергера и А.Д.Наумова). Материалы 2-й Международной конференции). 18-22 июля 2007 г., м. Картеш. – СПб, ЗИН РАН, 2007. с 28-29
Куликова Т.П, Видовой состав зоопланктона внутренних водоемов Карелии // Тр. КарНЦРАН. Биогеография Карелии. Серия Б. Вып. 2. Петрозаводск, 2001. С. 133–157
Мордухай-Болтовской Ф.Д., Ривьер И.К. Хищные ветвистоусые Podonidae, Polyphemidae, Cercopagidae и Leptodoridae фауны мира. - Л.: Наука, 1987. -182 с.
Полякова Н.В., Старков А.И., Кошелева А.Н., Сухих Н.М. 2006. Зоопланктон наскальных ванн островов Керетского архипелага Кандалакшского залива Белого моря. // Материалы Х научной конференции ББС
им. Перцова, М. С. 78-80.
Стогов И.А., Мовчан Е.А., Полякова Н.В., Сухопарова Е.Ю. 1996. Физико-химические и биологические характеристики наскальных ванн островов Керетского архипелага Кандалакшского залива Белого моря в 1990-1994 гг. Вестник СПбГУ, серия 3, вып. 2, N 10, с.17-23
Стогов, И.А., Полякова, Н.В., Старков, A.И, Мовчан Е.А. 2010. Планктонные беспозновоночные водоемов побережья Белого моря // Материалы XI Всероссийской конференции с международным участием «Проблемы изучения, рационального использования и охраны природных ресурсов Белого моря», 9–11 ноября 2010 г. – С.-Петербург, под ред. А.Д. Наумова, А.П. Алексеева и А.А. Сухотина,
— с. 179—182, ЗИН РАН, С.-Петербург
Царёв И.Л., Волкова П.А., Глаголев С.М. 2011. Изучение морфологической изменчивости
Daphnia longispina (Cladocera, Crustacea) на острове Асафий (Кандалакшский залив Белого моря) методами классической и геометрической морфометрии // Зоологический журнал. Т. 90 (1). С. 109-114
Altermatt F., Pajunen V.I., Ebert D. 2009. Desiccation of rock pool habitats and its influence on population persistence in a
Daphnia metacommunity. PLoS ONE, v.4, N 3, e4703
Armstrong R.A. 1984. The influence of bird droppings and uric acid on the radial growth of five species of saxicolous lichens // Environmental and Experimental Botany, V.24, № 1, pp 95-99
Bengtsson J. Life histories and interspecific competition between three Daphnia species in rockpools. // J.Anim.Ecol., 1986, V 55, N 2, p. 641-655.
Bengtsson J. Life histories, interspecific competition and regional distribution of three rockpool
Daphnia species. // Acta univ.upsal. Compr.summ.Uppsala Diss.Fac.Sci., 1988, N 142, p. 1-32.
Bengtsson J. Interspecific competition increases local extinction rate in a metapopulation system. 1989. Nature, v.340, N 6236, p.713-715
Brendonck L., Jocque M., Hulsmans A., Vanschoenwinkel B. 2010. Pools ‘on the rocks’: freshwater rock pools as model system in ecological and evolutionary research. Limnetica, v.21, N1, p.25-40.
Bohonak A.J., Jenkins D.G. 2003. Ecological and evolutionary significance of dispersal by freshwater invertebrates. Ecology Letters, v.6, N 8, p.783-796
Colwell, R. K., C. X. Mao, & J. Chang. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 85, 2717-2727
Colwell, R.K., 2006. EstimateS: Statistical estimation of species richness and shared species from samples. Version 8. Persistent URL
Dumont H.J., Segers H. 1996. Estimating lacustrine zooplankton species richness and complementarity. Hydrobiologia 341 (2), p. 125-132
Fryer G. 1980. Acidity and species diversity in freshwater crustacean faunas. Freshwater Biology, V.10, N.1, pp 41-45;
Fryer G. 1985. Crustacean diversity in relation to the size of water bodies: some facts and problems. Freshwater Biology, V.15, N 3, pp 347-361
Ghilarov A.M., The zooplankton of arctic rock pools // Oikos, 1967, N 18(1), p. 83-85.
Gotelli N.J., Colwell R.K. 2001. Quantifying biodiversity: procedures and pitfalls in the measurment and comparison of species richness. Ecology letters 4, p. 379-391
Haag C.R., Riek M., Hottinger J.W., Pajunen V.I., Ebert D. 2005. Genetic diversity and genetic differentiation in
Daphnia metapopulations with subpopulations of known age. Genetics, v.170, N4, 1809-1820
Koivisto S. Is Daphnia magna an ecologically representative zooplankton species in toxicity tests? 1995, Environmental Pollution, Volume 90, Issue 2, Pages 263-267
Lieder U. 1983 Revision of the genus Bosmina Baird 1845 (Crustacea, Cladocera). Int Revue ges Hydrobiol, 68(1), p. 121-139
MacArthur, R. H. and Wilson, E. O. 1967. The Theory of Island Biogeography. Princeton, N.J.: Princeton University Press.
Manny B.A., Johnson W.C. & Wetzel R.G. 1994. Nutrient additions by waterfowl to lakes and reservoirs: predicting their effects on productivity and quality. Hydrobiologia 279/280: 133-147
Nilssen J.P., Hobaek A., Petrusek A., Skage M., Restoring Daphnia lacustris G.O. Sars, 1862 (Crustacea, Anomopoda) – a cryptic species in the
Daphnia longispina group. 2007 Hydrobiologia 594 (1), p. 5-17
Pajunen V.I., Pajunen I., 2003. Long-term dynamics in rock pool Daphnia metapopulations. // Ecography N 26, p. 731-738.
Pajunen V.I., Pajunen I. 2007. Habitat characteristics contributing to local occupancy and habitat use in rock pool
Daphnia metapopulations. Hydrobiologia 592 (1), p. 291-302
R Development Core Team., 2004. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing [Electronic resource]. http: // www.R_project.org
Ranta E. Niche of Daphnia species in rock pools. Arch.Hydrobiol., 1979, N 87, p. 205-223
Ranta E. Animal communities in rock pools. Ann.Zool.Fennici, 1982, N 19, p. 337-347
Sacherov V., Havel L. 1997. Subgenic differences in head shield and ephippia ultrastructure within the genus
Bosmina Baird, 1845 (Crustacea, Cladocera). East 360, p. 13-23
Srivastava D.S., Kolasa J., Bengtsson J., Gonzalez A., Lowler S.P., Miller T.E., Munguia P., Romanuk T., Schneider D.C., Trzcinski M.K. 2004. Are natural microcosms useful model systems for ecology? Trends in Ecology and Evolution, v.19, N7, p.379-384