Насекомоядность рассматривается как способ получения дополнительных минеральных веществ, в основном азота и фосфора, на бедных минеральными веществами почвах (Chandler, Anderson, 1976; Karlsson, Carlsson, 1984). Некоторые авторы (Ellison, Gotelli, 2001) отмечают, что потребность в органическом азоте у насекомоядных растений уменьшается по мере упрощения строения ловчих структур, и у представителей рода Drosera невелика. Роль насекомоядности для растения может варьировать в зависимости от структуры сообщества и содержания в почве различных питательных веществ (Kraft, Handel, 1991). По мнению R. Dore Swamy и H.J. Maham Ram (1969) и J. D. C. Small с соавторами (1977), насекомоядность у росянок факультативна и, кроме того, слабо влияет на их рост в естественных условиях (Stewart, Nilsen, 1992).
Поведение листьев представителей рода Drosera при поимке своих жертв было подробно описано более чем 100 лет назад (Дарвин, 1908; Hooker, 1916). Листья росянок имеют короткие железистые ловчие волоски в центре листовой пластинки и более длинные (краевые) волоски по ее периферии.
При попадании жертвы на листовую пластинку все волоски изгибаются навстречу добыче и обволакивают ее слизью, содержащей пищеварительные ферменты. Наблюдаются как быстрые движения железистых волосков при контакте с животным (через 10-30 сек после контакта), так и медленные движения волосков, не контактировавших непосредственно с животным, через несколько часов после его попадания на листовую пластинку (Hooker, 1916; Bopp, Weber, 1981). B. Juniper с соавторами (1989) считают, что у Drosera слизь выделяется только в ответ на попадание жертвы на листовую пластинку. Наиболее интенсивная секреция слизи у D. rotundifolia наблюдается на вторые сутки после попадания на лист добычи (Муравник, 2002). После выделения слизи край листа медленно загибается и закрывает перевариваемую добычу.

По окончании переваривания лист разворачивается, волоски выпрямляются, и слизь высыхает (Hooker, 1916).
Однако такой механизм ловли животных, по-видимому, не очень надежен: неоднократно было отмечено произвольное освобождение насекомых из листовых ловушек D. filiformis (Gibson, 1991). Кроме того, поведение ловчих листьев насекомоядных растений зависит не только от числа и состояния насекомых, находящихся на листовой пластинке. Установлено, например, что секреторная активность железистых волосков у D. capillaris максимальна при высокой относительной влажности воздуха и низкой интенсивности света (Gomez, 1998).
Н.В. Осташева (2002) выяснила, что для листьев D. rotundifolia характерны ритмичные движения, не связанные с улавливанием насекомых. На севере Карелии период движения листьев сильно варьировал и равнялся примерно 15 ч, а в Вологодской области -- 12 ч.
К сожалению, большая часть наблюдений за поведением ловчих листьев насекомоядных растений сделана в лабораторных условиях, что не позволяет учитывать естественные колебания числа доступных насекомых, а также влияние погодных факторов на поведение листьев. Ранее мы неоднократно исследовали различные аспекты биологии D. rotundifolia в природных условиях (Волкова и др., 2001; Волкова, 2002a; Волкова, 2002b; Волкова, Шипунов, 2002; Волкова и др., 2002; Волкова и др., 2003). Целью настоящей работы является изучение поведения ловчих листьев D. rotundifolia в естественных условиях и выявление влияния на него погодных условий и пойманных насекомых.
Наблюдения велись: (1) за двумя растениями D. rotundifolia в Лоухском районе республики Карелия, мыс Иванов Наволок (66 град. 20' с.ш., 33 град. 20' в.д.) с 25 по 27 июля 2000 года -- серия 1, и (2) за двумя растениями D. rotundifolia в Вышневолоцком районе Тверской области, западный берег озера Ольшево (58 град. 15' с.ш., 34 град. 30' в.д.) с 20 по 23 июня 2002 года -- серия 2.
Продолжительность светового дня 29 июля в Лоухском районе составляла 19 ч 17 мин при отсутствии темного времени суток, 29 июня в Вышневолоцком районе -- 18 ч 17 мин (данные получены при помощи компьютерной астрономической программы XEphem -- Downey, 2000). Растения были выбраны случайным образом из типичных для данных природных условий популяций. Непрерывные наблюдения за каждой парой исследуемых растений, находившихся в естественных условиях, продолжались в течение 72 часов. В течение всего срока наблюдений для каждого из 16 исследованных ловчих листьев D. rotundifolia (табл. 2) один раз в 40 минут в 2001 году и один раз в 30 минут в 2002 году визуально оценивали форму листовой пластинки, степень ее увлажненности липкой слизью и долю загнутых краевых железистых волосков (табл. 1), а также подсчитывали количество насекомых, находящихся на листовой пластинке.
Таблица 1. Критерии визуальной оценки состояния листовой пластинки
| баллы | степень увлажненности листовой пластинки ("wet") | форма листовой пластинки ("shape") | доля загнутых краевых железистых волосков от общего числа железистых волосков ("tent") |
| 0 | слабая (лист практически сухой) | ... | загнутых волосков практически нет |
| 1 | средняя | практически плоская | меньшая часть |
| 2 | сильная (слизь собирается в капли) | вогнутая | около половины |
| 3 | ... | завернутая | большая часть |
| 4 | ... | ... | практически все волоски загнуты |
Таблица 2. Распределение исследованных листьев по растениям
| номер серии | число исследованных листьев (шт.) | |
| на первом растении | на втором растении | |
| 1 | 5 | 4 |
| 2 | 3 | 4 |
В течение наблюдений 2002 года один раз в два часа измеряли атмосферное давление при помощи бытового барометра, температуру и относительную влажность воздуха при помощи аспирационного психрометра Ассмана.
В ходе обработки данных для каждого исследованного растения были вычислены также "средняя форма листовой пластинки", "средняя степень увлажненности листовой пластинки" и "средняя доля загнутых краевых волосков" путем усреднения значений соответствующих характеристик для всех листьев данного растения за одно наблюдение. Для выявления связи между исследованными характеристиками листовой пластинки, погодными условиями и количеством насекомых на листовой пластинке был проведен рассчет непараметрических коэффициентов корреляции Спирмена (табл. 3, 5). Для вычислений использовался пакет программ STATISTICA (версия STATISTICA for Windows; StatSoft, Inc., 1999).
Таблица 3. Непараметрические коэффициенты корреляции Спирмена между характеристиками листовых пластинок в серии 1 (N=81, включены статистически достоверные коэффициенты корреляции с p<0,01 и абсолютным значением >0,4)
Часть 1. Взаимосвязь характеристик состояния листовой пластинки отдельных листьев
| коррелирующие параметры | коэффициент корреляции |
| tent13 & shape13 | 0,63 |
| tent11 & shape14 | -0,62 |
| tent11 & tent15 | 0,54 |
| tent11 & tent21 | 0,53 |
| shape14 & tent22 | -0,52 |
| tent11 & tent23 | 0,48 |
| tent23 & wet23 | -0,48 |
| shape14 & tent15 | -0,47 |
| shape14 & wet24 | 0,46 |
| tent13 & tent21 | 0,46 |
| tent11 & tent22 | 0,43 |
| tent15 & tent21 | 0,43 |
| shape23 & shape24 | 0,42 |
| shape21 & shape23 | 0,42 |
| wet11 & wet12 | 0,42 |
| wet23 & wet24 | 0,40 |
| shape14 & tent23 | -0,40 |
Часть 2. Связь характеристик состояния листовой пластинки отдельных листьев с усредненными по всем листьям растения характеристиками состояния листовой пластинки
| коррелирующие параметры | коэффициент корреляции |
| wet2 av & wet23 | 0,90 |
| shape2 av & shape23 | 0,82 |
| wet1 av & wet12 | 0,76 |
| tent2 av & tent22 | 0,76 |
| tent1 av & tent15 | 0,75 |
| wet2 av & wet24 | 0,75 |
| wet1 av & wet15 | 0,68 |
| tent1 av & tent11 | 0,67 |
| tent2 av & tent23 | 0,66 |
| tent2 av & tent11 | 0,66 |
| shape2 av & shape24 | 0,64 |
| wet1 av & wet11 | 0,63 |
| shape1 av & shape11 | 0,63 |
| tent2 av & shape14 | -0,61 |
| tent2 av & tent21 | 0,60 |
| shape2 av & shape21 | 0,58 |
| tent1 av & tent21 | 0,58 |
| shape1 av & shape13 | 0,55 |
| tent1 av & tent12 | 0,54 |
| shape1 av & shape12 | 0,51 |
| tent2 av & tent15 | 0,51 |
| shape1 av & shape14 | 0,50 |
| shape2 av & shape22 | 0,49 |
| wet2 av & tent23 | -0,49 |
| tent2 av & wet23 | -0,47 |
| wet2 av & shape14 | 0,46 |
| tent1 av & shape14 | -0,46 |
| wet1 av & wet13 | 0,44 |
| tent2 av & tent24 | 0,43 |
| tent1 av & tent14 | 0,42 |
| shape1 av & tent13 | 0,41 |
| tent1 av & tent22 | 0,40 |
Часть 3. Взаимосвязь усредненных по всем листьям растения характеристик состояния листовой пластинки (все коэффициенты)
| коррелирующие параметры | коэффициент корреляции |
| tent1 av & tent2 av | 0,59 |
| wet2 av & tent2 av | -0,54 |
| tent1 av & wet2 av | -0,33 |
| wet2 av & shape2 av | -0,25 |
Условные обозначения:
"wetNM" -- степень увлажненности листовой пластинки M-ного листа N-ного растения (см. также табл. 1)
"wetN av" -- усредненная ("av") по всем листьям N-ного растения степень увлажненности листовой пластинки
Таблица 5. Непараметрические коэффициенты корреляции Спирмена между характеристиками листовых пластинок в серии 2 (N=144, включены статистически достоверные коэффициенты корреляции с p<0,01 и абсолютным значением >0,4)
Часть 1. Взаимосвязь характеристик листовой пластинки отдельных листьев
| коррелирующие параметры | коэффициент корреляции |
| wet11 & wet13 | 0,82 |
| wet21 & wet22 | 0,73 |
| wet21 & wet11 | 0,73 |
| shape11 & tent11 | 0,69 |
| wet23 & wet12 | 0,69 |
| wet22 & wet13 | 0,68 |
| wet22 & wet11 | 0,68 |
| wet21 & wet13 | 0,67 |
| tent11 & tent13 | 0,66 |
| wet11 & wet12 | 0,65 |
| tent22 & tent24 | 0,64 |
| wet21 & wet23 | 0,64 |
| wet21 & wet24 | 0,64 |
| wet12 & wet13 | 0,63 |
| wet21 & wet12 | 0,63 |
| wet22 & wet24 | 0,62 |
| tent22 & tent23 | 0,61 |
| tent24 & tent13 | 0,60 |
| wet24 & wet12 | 0,59 |
| wet23 & wet24 | 0,58 |
| wet22 & wet23 | 0,58 |
| shape24 & shape13 | 0,57 |
| shape22 & shape11 | 0,57 |
| wet24 & wet13 | 0,56 |
| tent21 & tent23 | 0,55 |
| shape22 & shape24 | 0,55 |
| shape21 & shape11 | 0,54 |
| tent22 & tent13 | 0,54 |
| tent22 & tent11 | 0,54 |
| shape21 & shape22 | 0,53 |
| tent23 & tent24 | 0,53 |
| wet24 & wet11 | 0,53 |
| shape12 & tent12 | 0,52 |
| shape13 & tent13 | 0,50 |
| wet22 & wet12 | 0,50 |
| tent21 & tent22 | 0,50 |
| shape24 & shape11 | 0,50 |
| wet23 & wet13 | 0,49 |
| tent21 & tent11 | 0,49 |
| wet23 & wet11 | 0,48 |
| tent24 & tent11 | 0,48 |
| tent21 & tent13 | 0,48 |
| tent23 & tent11 | 0,47 |
| tent23 & tent13 | 0,46 |
| shape11 & shape13 | 0,46 |
| shape21 & shape24 | 0,45 |
| shape22 & shape13 | 0,44 |
| shape21 & tent11 | 0,44 |
| shape21 & shape13 | 0,43 |
| shape22 & shape12 | 0,41 |
| tent22 & shape11 | 0,41 |
| tent24 & tent12 | 0,40 |
| shape22 & tent11 | 0,40 |
Часть 2. Связь усредненных по всем листьям растения характеристик состояния листовой пластинки с характеристиками состояния отдельных листьев
| коррелирующие параметры | коэффициент корреляции |
| wet12 & wet1 av | 0,89 |
| wet11 & wet1 av | 0,88 |
| wet21 & wet2 av | 0,88 |
| tent22 & tent2 av | 0,87 |
| wet13 & wet1 av | 0,86 |
| tent11 & tent1 av | 0,84 |
| wet23 & wet2 av | 0,84 |
| wet22 & wet2 av | 0,84 |
| shape11 & shape1 av | 0,83 |
| tent24 & tent2 av | 0,82 |
| wet24 & wet2 av | 0,81 |
| tent23 & tent2 av | 0,81 |
| tent13 & tent1 av | 0,80 |
| shape22 & shape2 av | 0,80 |
| shape24 & shape2 av | 0,76 |
| shape21 & shape2 av | 0,75 |
| shape12 & shape1 av | 0,74 |
| wet21 & wet1 av | 0,74 |
| shape13 & shape1 av | 0,73 |
| tent12 & tent1 av | 0,72 |
| wet12 & wet2 av | 0,71 |
| wet11 & wet2 av | 0,70 |
| wet13 & wet2 av | 0,68 |
| tent21 & tent2 av | 0,67 |
| wet23 & wet1 av | 0,65 |
| wet22 & wet1 av | 0,64 |
| tent13 & tent2 av | 0,64 |
| wet24 & wet1 av | 0,62 |
| shape22 & shape1 av | 0,61 |
| tent11 & tent2 av | 0,61 |
| tent11 & shape1 av | 0,61 |
| shape11 & shape2 av | 0,59 |
| shape24 & shape1 av | 0,59 |
| tent24 & tent1 av | 0,59 |
| shape23 & shape2 av | 0,59 |
| shape13 & shape2 av | 0,56 |
| shape11 & tent1 av | 0,54 |
| shape21 & shape1 av | 0,53 |
| tent22 & tent1 av | 0,53 |
| tent21 & tent1 av | 0,50 |
| tent13 & shape1 av | 0,47 |
| tent23 & tent1 av | 0,43 |
| shape12 & tent1 av | 0,43 |
| shape21 & tent1 av | 0,43 |
| shape21 & tent2 av | 0,41 |
| shape13 & tent1 av | 0,41 |
| shape22 & tent1 av | 0,40 |
| tent22 & shape1 av | 0,40 |
Часть 3. Связь характеристик состояния листовой пластинки отдельных листьев с регистрируемыми погодными условиями
| коррелирующие параметры | коэффициент корреляции |
| относительная влажность воздуха & wet13 | 0,62 |
| относительная влажность воздуха & wet11 | 0,61 |
| относительная влажность воздуха & wet12 | 0,60 |
| относительная влажность воздуха & wet22 | 0,59 |
| температура воздуха & wet11 | -0,56 |
| температура воздуха & wet12 | -0,55 |
| относительная влажность воздуха & wet21 | 0,54 |
| температура воздуха & wet13 | -0,54 |
| атмосферное давление & shape11 | 0,53 |
| атмосферное давление & tent11 | 0,52 |
| относительная влажность воздуха & wet23 | 0,49 |
| относительная влажность воздуха & wet24 | 0,48 |
| температура воздуха & wet22 | -0,47 |
| температура воздуха & wet21 | -0,42 |
| атмосферное давление & tent21 | 0,40 |
Часть 4. Взаимосвязь усредненных по всем листьям растения характеристик состояния листовой пластинки (все коэффициенты)
| коррелирующие параметры | коэффициент корреляции |
| wet2 av & wet1 av | 0,78 |
| tent2 av & tent1 av | 0,65 |
| shape2 av & shape1 av | 0,64 |
| shape1 av & tent1 av | 0,61 |
| tent2 av & shape1 av | 0,41 |
| shape2 av & tent1 av | 0,36 |
| shape2 av & tent2 av | 0,34 |
| wet2 av & shape2 av | 0,24 |
| shape2 av & wet1 av | 0,24 |
Часть 5. Связь усредненных по всем листьям растения характеристик состояния листовой пластинки с регистрируемыми погодными условиями (все коэффициенты)
| коррелирующие параметры | коэффициент корреляции |
| относительная влажность воздуха & wet1 av | 0,68 |
| относительная влажность воздуха & wet2 av | 0,61 |
| температура воздуха & wet1 av | -0,61 |
| атмосферное давление & tent1 av | 0,56 |
| атмосферное давление & shape1 av | 0,48 |
| температура воздуха & wet2 av | -0,46 |
| атмосферное давление & tent2 av | 0,41 |
| температура воздуха & shape1 av | 0,23 |
| атмосферное давление & wet2 av | -0,23 |
| относительная влажность воздуха & tent2 av | 0,20 |
| атмосферное давление & shape2 av | 0,20 |
Условные обозначения:
см. условные обозначения к табл. 3
Мы не отмечали реакции листовой пластинки на насекомых в течение часа после попадания их на листовую пластинку (табл. 4).
Таблица 4. Поведение листьев в серии 1
| номер листа | характеристика состояния листовой пластинки | изменение состояния после попадания насекомого на лист через ... | изменение состояния без насекомых на листе | изменение состояния при постоянном количестве насекомых | отличное от описанного в литературе поведения при наличии насекомых | ||
| менее 1 ч | 1--2 ч | 3 ч | |||||
| первый лист первого растения | увлажненность | ... | ... | ... | + | ... | ... |
| форма | ... | ... | ... | + | ... | ... | |
| положение краевых волосков | ... | ... | ... | + | ... | ... | |
| второй лист первого растения | увлажненность | -- | -- | + | ... | + | + |
| форма | -- | -- | -- | ... | + | + | |
| положение краевых волосков | -- | + | -- | ... | + | + | |
| третий лист первого растения | увлажненность | -- | -- | -- | + | + | + |
| форма | -- | -- | + | + | + | -- | |
| положение краевых волосков | -- | -- | + | + | -- | -- | |
| четвертый лист первого растения | увлажненность | -- | -- | -- | -- | -- | + |
| форма | -- | -- | -- | -- | + | + | |
| положение краевых волосков | -- | -- | -- | + | + | + | |
| пятый лист первого растения | увлажненность | ... | ... | ... | ... | + | + |
| форма | ... | ... | ... | ... | + | -- | |
| положение краевых волосков | ... | ... | ... | ... | + | -- | |
| первый лист второго растения | увлажненность | ... | ... | ... | ... | -- | + |
| форма | ... | ... | ... | ... | + | -- | |
| положение краевых волосков | ... | ... | ... | ... | -- | -- | |
| второй лист второго растения | увлажненность | -- | -- | -- | -- | -- | + |
| форма | -- | + | -- | + | + | -- | |
| положение краевых волосков | -- | + | -- | + | + | + | |
| третий лист второго растения | увлажненность | -- | -- | -- | + | + | -- |
| форма | -- | -- | -- | + | + | -- | |
| положение краевых волосков | -- | -- | + | + | + | -- | |
| четвертый лист второго растения | увлажненность | -- | -- | -- | + | + | + |
| форма | -- | -- | -- | + | + | -- | |
| положение краевых волосков | -- | + | + | + | + | + | |
| доля листьев с указанными изменениями, % | увлажненность | 0 | 0 | 17 | 67 | 63 | 88 |
| форма | 0 | 17 | 17 | 83 | 100 | 25 | |
| положение краевых волосков | 0 | 50 | 50 | 100 | 75 | 50 | |
Условные обозначения:
"..."' -- указанный режим нахождения насекомых на листовой пластинки для данного листа не отмечен,
"+" -- наблюдались указанные изменения данной характеристики состояния листовой пластинки,
"--" -- указанные изменения данной характеристики состояния листовой пластинки не наблюдались,
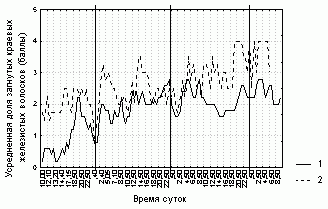
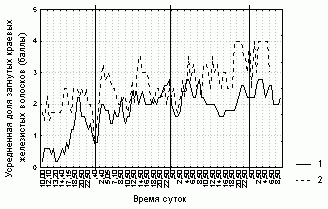
Только через 2-3 часа практически у всех исследованых листьев увеличивалась доля загнутых краевых волосков. Изменения степени увлажненности листовой пластинки и ее формы в ответ на попадание насекомого практически не наблюдалось. У всех исследованных листьев в отсутствие насекомых или при наличии постоянного числа непереваренных насекомых наблюдались частые изменения состояния. При наличии непереваренного насекомого практически все исследованные листья периодически высыхали, у половины исследованых листьев не были загнуты краевые волоски, а у четверти листовая пластинка не была завернута. Таким образом, нами не было обнаружено достоверной связи между различными исследованными характеристиками состояния листовой пластинки практически для всех исследованных листьев (рис. 1-3, табл. 3).
Рисунок 1. Изменение степени увлажненности листовой пластинки D. rotundifolia в районе Полярного Круга
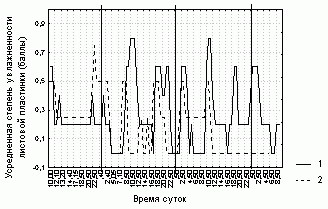
Условные обозначения:
Баллы состояния листовой пластинки см. табл. 1.
Вертикальными сплошными линиями отмечено 00:00
часов каждых суток наблюдения.
1 -- первое исследуемое растение D. rotundifolia,
2 -- второе исследуемое растение
Рисунок 2. Изменение формы листовой пластинки D. rotundifolia в районе Полярного Круга
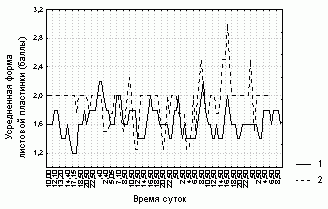
Условные обозначения:
см. условные обозначения к рис. 1
Рисунок 3. Изменение доли загнутых краевых железистых волосков на листовой пластинке D. rotundifolia в районе Полярного Круга
Условные обозначения:
см. условные обозначения к рис. 1
Степень увлажненности листовой пластинки, ее форма и доля загнутых краевых волосков у каждого листа не очень четко (R=0,4-0,7, p=0) связаны со значениями этих характеристик, усредненных по всем листьям растения (табл. 3). Отсюда следует, что усредненные характеристики состояния листовой пластинки способны описывать общие тенденции поведения каждого листа данного растения. Более того, изменения различных характеристик листовой пластинки на разных растениях согласованы (рис. 1-3, табл. 3).
Примерно у половины листьев наблюдалась реакция на пойманное насекомое менее чем через час после попадания насекомого на листовую пластинку (табл. 6).
Таблица 6. Поведение листьев в серии 2
| номер листа | характеристика состояния листовой пластинки | изменение состояния после попадания насекомого на лист через ... | изменение состояния без насекомых на листе | изменение состояния при постоянном количестве насекомых | отличное от описанного в литературе поведения при наличии насекомых | ||
| менее 1 ч | 1--2 ч | 3 ч | |||||
| первый лист первого растения | увлажненность | -- | -- | + | + | + | + |
| форма | + | -- | -- | + | -- | -- | |
| положение краевых волосков | -- | -- | -- | + | + | -- | |
| второй лист первого растения | увлажненность | -- | -- | + | + | + | + |
| форма | -- | + | + | + | + | -- | |
| положение краевых волосков | + | + | + | + | + | + | |
| третий лист первого растения | увлажненность | + | -- | -- | + | ... | + |
| форма | + | -- | -- | + | ... | -- | |
| положение краевых волосков | + | -- | -- | + | ... | + | |
| первый лист второго растения | увлажненность | + | -- | -- | + | -- | -- |
| форма | + | -- | -- | + | -- | -- | |
| положение краевых волосков | + | -- | -- | + | -- | -- | |
| второй лист второго растения | увлажненность | -- | -- | + | ... | + | + |
| форма | -- | -- | + | ... | + | + | |
| положение краевых волосков | + | -- | -- | ... | + | + | |
| третий лист второго растения | увлажненность | + | -- | + | + | + | + |
| форма | + | + | -- | + | + | + | |
| положение краевых волосков | -- | + | + | + | + | + | |
| четвертый лист второго растения | увлажненность | -- | -- | + | ... | + | + |
| форма | -- | + | -- | ... | + | -- | |
| положение краевых волосков | + | -- | -- | ... | + | + | |
| доля листьев с указанными изменениями, % | увлажненность | 43 | 0 | 70 | 100 | 83 | 86 |
| форма | 57 | 43 | 29 | 100 | 67 | 29 | |
| положение краевых волосков | 71 | 29 | 29 | 100 | 83 | 71 | |
Условные обозначения: см. условные обозначения к табл. 4
У 40% листьев за этот период времени происходило увлажнение листовой пластинки, у 50% листьев листовая пластинка заворачивалась, а у 70% листьев увеличивалась доля загнутых краевых железистых волосков. Практически у всех листьев, не изменивших свое состояние менее чем через час после попадания на них насекомых, наблюдалось заворачивание и увлажнение листовой пластинки, а также загибание краевых волосков в течение двух-трех часов с момента попадания насекомого на листовую пластинку. При этом, с одной стороны, степень увлажненности листовой пластинки и степень ее завернутости у 60-70% от общего числа исследованных листьев была прямо связана с числом насекомых на листовой пластинке. С другой стороны, у всех листьев в отсутствии насекомых на листовой пластинке наблюдались не имеющие четкой периодичности изменения степени ее увлажненности и формы продолжительностью 1-6 часов, а также доли загнутых краевых волосков.
Такое поведение было отмечено и для большинства (70%-80% от общего числа) исследованных листьев в присутствии на их листовой пластинке постоянного числа непереваренных насекомых.
Часто наблюдалось отличное от описанного в литературе поведение листа при наличии на его пластинке недавно пойманного насекомого. Так, практически все исследованные листья не выделяли слизи при попадании на них насекомых, а краевые волоски 70% листьев не были загнутыми при нахождении на листовой пластинке непереваренного насекомого.
Степень загнутости листовой пластинки всех исследованных листьев прямо связана (R=0,4-0,6, p<0,001) с долей загнутых краевых железистых волосков на этой листовой пластинке (табл. 5). Нами не было обнаружено достоверной связи между степенью увлажненности листовой пластинки и ее формой, а также долей загнутых краевых волосков на листе (рис. 4-6, табл. 5).
Рисунок 4. Изменение степени увлажненности листовой пластинки D. rotundifolia в умеренной зоне Европейской России
Условные обозначения:
см. условные обозначения к рис. 1
Рисунок 5. Изменение формы листовой пластинки D. rotundifolia в умеренной зоне Европейской России
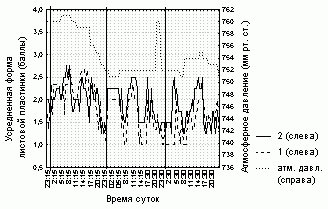
Условные обозначения:
см. условные обозначения к рис. 1
Рисунок 6. Изменение доли загнутых краевых железистых волосков на листовой пластинке D. rotundifolia в умеренной зоне Европейской России
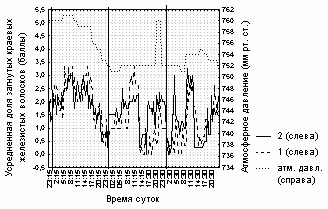
Условные обозначения:
см. условные обозначения к рис. 1
Аналогично серии 1, степень увлажненности листовой пластинки, ее форма и доля загнутых краевых волосков для каждого листа в серии 2 достоверно коррелируют (R=0,7-0,9, p=0) со значениями этих характеристик, усредненных по всем листьям растения (табл. 5). Состояние листовой пластинки изменялось синхронно практически у всех исследованных листьев, вне зависимости от того, какому растению они принадлежат (рис. 4-6, табл. 5).
Степень увлажненности листовой пластинки у всех исследованных листьев прямо зависит (R=0,5-0,7, p=0) от относительной влажности воздуха (рис. 4, табл. 5). Для всех исследованных листьев также отмечена более слабая положительная зависимость (R=0,2-0,5, p<0,05) доли загнутых краевых волосков и степени завернутости листовой пластинки от атмосферного давления (рис. 5, 6, табл. 5). При этом зависимости числа пойманных насекомых от погодных условий выявлено не было.
В период наблюдений температура воздуха в среднем оставалась постоянной и составляла 17 град. C. Суточные колебания температуры воздуха были хорошо выражены: температура достигала своего минимального значения в 5-6 утра, опускаясь до 9 град. С, и своего максимального значения в 14-15 дня, поднимаясь до 26,4 град. С. Заметного влияния температуры воздуха на характеристики листовой пластинки не было обнаружено. Наблюдаемая в некоторых случаях слабая зависимость характеристик листовой пластинки от температуры воздуха, по-видимому, опосредована довольно тесной отрицательной связью температуры воздуха с его относительной влажностью.
Наши наблюдения показывают, что различные характеристики листовой пластинки D. rotundifolia как при типичном для средней полосы фоторежиме, так и в условиях полярного дня изменяются независимо друг от друга и от наличия насекомых на листе. Интересно отметить, что в серии 2 наблюдались более согласованные изменения различных характеристик листовой пластинки, чем в серии 1. Кроме того, в серии 2 реакция листа на пойманное насекомое была выражена более четко, чем у росянок в серии 1. Эти различия, вероятно, связаны с более жесткими условиями произрастания D. rotundifolia в районе Полярного Круга по сравнению со средней полосой Европейской России, что могло негативно отразиться на активности "охотничьего поведения" росянок из северных популяций.
Синхронность изменения различных характеристик состояния листовой пластинки у листьев различных растений также косвенно свидетельствует об отсутствии существенной зависимости поведения ловчих листьев от наличия на них насекомых. Это подтверждается наблюдениями Н.В. Осташевой (2002), выполненными в полевых условиях, и не согласуется с традиционными взглядами на поведение ловчих листьев D. rotundifolia, как правило, основанными на лабораторных экспериментах (Дарвин, 1908; Hooker, 1916; Bopp, Weber, 1981, Juniper et al., 1989, Муравник, 2002).
По-видимому, степень увлажненности листовой пластинки насекомоядных растений как и листьев всех прочих растений не может не зависеть от относительной влажности воздуха, что подтверждают данные L.D. Gomez (1998) для D. capillaris. Возможно, в ответ на попадание насекомых на листовую пластинку меняется лишь химический состав ловчей слизи, тогда как ее количество остается практически постоянным. Наблюдаемая зависимость формы листовой пластинки и доли загнутых волосков от атмосферного давления объясняется, по-видимому, обусловленностью движений листовой пластинки и железистых волосков изменениями внутриклеточного давления. Проводимые, как правило, в лабораторных условиях наблюдения за поведением ловчих листьев (Дарвин, 1908; Hooker, 1916; Муравник, 2002) не могли учитывать влияние погодных факторов на состояние листовой пластинки, которое в естественных условиях, похоже, превышает эффект от пойманных насекомых.
Нам представляется, что изменение состояния листовой пластинки у D. rotundifolia, возможно, носит случайный характер и лишь корректируется внешними факторами, такими как относительная влажность воздуха, атмосферное давление и наличие насекомых на листовой пластинке. Нами не было обнаружено четкой ритмичности изменения состояния листовой пластинки как в условиях естественного для средней полосы фоторежима, так и в условиях полярного дня, что не согласуется с мнением Н.В. Осташевой (2002). Наблюдаемое увлажнение листовой пластинки, ее заворачивание, а также загибание краевых волосков через некоторое время после попадания насекомого на листовую пластинку можно, по-видимому, рассматривать как совпадение. Такая ситуация не уникальна, поскольку активная реакция ловчих структур на пойманное насекомое не является обязательной характеристикой насекомоядных растений. Например, у близкого Droseraceae семейства Nepenthaceae известны кувшинообразные ловушки, не удерживающие активно добычу (Ellison, Gotelli, 2001).
В качестве дополнительных аргументов в пользу отсутствия четкой реакции ловчего листа на насекомое могут рассматриваться не только случаи произвольного освобождения насекомых из листовой ловушки (Gibson, 1991; Zamora, 1999), но и неоднократно наблюдавшиеся нами для D. rotundifolia и другими авторами для различных видов насекомоядных растений (Watson et al., 1982 -- D. erythrorhiza; Zamora, 1999 -- P. vallisnerifolia; Bingham, Cohen, 2001 -- D. rotundifolia) случаи клептопаразитизма.
Наши данные об отсутствии четкой зависимости состояния ловчих листьев от наличия на них насекомых у D. rotundifolia подтверждают развиваемую в последнее время гипотезу о факультативности насекомоядности для насекомоядных растений (Dore Swamy, Maham Ram, 1969; Small et al., 1977; Stewart, Nilsen, 1992; Ellison, Gotelli, 2001).
Сбор данных для настоящей работы проводился в ходе полевых практик Московской Гимназии на Юго-Западе, проходивших под руководством С. Глаголева. В полевых наблюдениях активнейшее участие принимали Е. Альтшулер, Т. Браславская, П. Бунтман, О. Васильева, Т. Волкова, Н. Горбунов, Я. Косенко, Д. Назаров, Е. Пескова, И. Покровский, В. Чава. Особая благодарность -- К. Марквичевой за иллюстрации. Ценные замечания при подготовке рукописи были сделаны Л.Е. Муравник и А.Н. Ивановой.
Волкова П.А., Кумскова Е.М., Назаров Д.Ю., Покровский И.Г. Взаимосвязь морфофизиологических характеристик, успеха ловли насекомых и условий обитания у различных видов насекомоядных растений // V Всероссийский популяционный семинар "Популяция, сообщество, эволюция". Часть 1. Казань: ЗАО "Новое издание", 2001. С. 14-17.
Волкова П.А., Кумскова Е.М., Шипунов А.Б. Зависимость морфофизиологических характеристик от успеха ловли насекомых и условий обитания у Drosera rotundifolia L., D. anglica Huds., D. obovata Mert. et Koch (Droseraceae) и Pinguicula vulgaris L. (Lentibulariaceae) // Бюллетень МОИП. отд. биол. 2003. Т. 108. Вып. 1. С. 72-78.
Волкова П. А. О морфологических отличиях трех видов росянки (Drosera L., Droseraceae) // Международная научная конференция по систематике высших растений, посвященная 70-летию со дня рождения чл.-корр. РАН, проф. В.Н. Тихомирова: Тез. докл. -- М.: Изд-во Центра охраны дикой природы, 2002a. С. 29-30.
Волкова П.А. Материалы по экологии Drosera rotundifolia L. на островах Керетского архипелага и губы Кив // III научная сессия морской биологической станции Санкт-Петербургского государственного университета: Тез. докл. -- СпБ., 2002b. С. 10-14.
Волкова П., Кумскова Е., Шипунов А. Особенности ловли насекомых у росянки круглолистной (Drosera rotundifolia L., Droseraceae) // Матеріали конференції молодих вчених-ботаніків України "Актуальні проблеми флористики, систематики, екології та збереження фіторізноманіття" (Львів, Івано-Франково, 6-10 серпня 2002 р.). - Львів, 2002. - С. 137-138.
Волкова П.А., Шипунов А.Б. Поведение ловчих листьев насекомоядного растения Drosera rotundifolia L. (Droseraceae). Экологическая ботаника: наука, образование, прикладные аспекты: Тез. докл. -- Сыктывкар, 2002. С. 61-62.
Дарвин Ч. Насекомоядные растения // Собр. соч. Ч. Дарвина. Т. 4. Ч. 2. М., 1908. С. 145-154.
Муравник Л.Е. Влияние химической стимуляции на ультраструктуру секреторных клеток железистых волосков двух видов Drosera // Физиол. раст. 2002. Т. 47, № 4. С. 614-623.
Осташева Н.В. Экотопическая приуроченность и изучение движений листьев видов Drosera. Курсовая работа, Москва, 2002. С. 11, 15.
Bingham R.A., Cohen N.D. Flowering, reproduction and kleptoparasitism in an extreme southern disjunct population of Drosera rotundifolia. Botany, 2001.
Bopp M., Weber I. Studies on the hormonal regulation of leaf blade movement of Drosera capensis L. Physiologia Plantarum, 1981, 53. 491-496.
Chandler G.E., Anderson J.W. Studies on the nutrition and growth of Drosera species with reference to the carnivorous habit // New Phytol. 1976. Vol. 76. P. 129-141.
Dore Swamy R., Maham Ram H.J. Studies on growth and flowering in axenic cultures of insectivorous plants. Phytomorphology, 19, 363.
Downey E.C. Xephem. Astronomy program. Version 3.2.2. 2000. [Electronic resource]. Mode of access: http://www.clearskyinstitute.com/xephem/.
Ellison A.M., Gotelli N.J. Evolutionary ecology of carnivorous plants // Trends in Ecol. and Evol. 2001. Vol. 16, N 11. P. 623-629.
Gibson T.C. Differential escape of insects from carnivorous plant traps // American Midland Naturalist. 1991. Vol. 125, N 1. P. 55-62.
Gomez L.D. Natural history and occurence of the "insectivorous plant" Drosera capillaris (Droseraceae) in Costa Rica // Revista de Biologia Tropical. 1998. Vol. 46, N 4. P. 1033-1037.
Hooker H.D. Physiological observations on Drosera rotundifolia // Bull. Torr. Bot. Club. 1916. Vol. 43, N 1. P. 1-27.
Juniper B.E., Robins R.J., Joel D.M. The carnivorous plants. Academic Press, London. 1989.
Karlsson P.S., Carlsson B. Why does Pinguicula vulgaris L. trap insects? // New Phytol. 1984. Vol. 97. P. 25-30.
Small J.G.C., Onraet A., Grierson D.S., Reynolds G. Studies on insect-free growth, development and nitrate-assimilating ensymes of Drosera aliciae hamet // New Phytol. 1977. Vol. 79. P. 127-133. StatSoft, Inc. 1999. STATISTICA for Windows [Computer Program Manual]. Tulsa, OK.
Stewart C.N., Nilsen E.T. Drosera rotundifolia growth and nutrition in a natural population with special reference to the significance of insectivory // Can. J. Bot. 1992. Vol. 70, N 7. P. 1409-1416.
Watson A.P., Mathiersseu J.N., Sprigett B.P. (1982). Arthropod associates and macronutrient status of the red-ink sundew (Drosera erythrorhiza Lindl.) Aust. J. Ecol. 7: 13-22.
Zamora R. Conditional outcomes of interactions: the pollinator-prey conflict of an insectictivorous plant // Ecology. 1999. Vol. 80, N 3. P. 786-795.
| Главная | Общая информация | Карты | Фото | Фольклор | Острова | Озера | Флора | Фауна |